Кинетика типичного В-клеточного ответа:
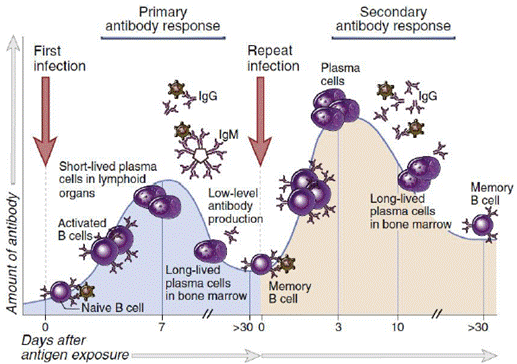
В ответ на АГ происходит активация и пролиферация В-клеток и продукция АТ плазматическими клетками. В первичном ответе больше растворимого IgM, меньше IgG. IgM дают хорошую защиту, и к тому моменту, как пройдут все реакции взаимодействия клеток в герминальных центрах, переключение изотипов на другие классы иммуноглобулинов и тд, к тому времени патоген будет нейтрализован IgM и переварен фагоцитами, иммунный ответ идет на спад. Клетки памяти после встречи переключили уже изотип на IgG (в результате рекомбинации в константной части), поэтому при повторной встрече с антигеном в основном продуцируются IgG, вторичный ответ сильнее и быстрее первичного – при повторной встрече за счет клеток памяти ответ более быстрый, специфичный и сильный. Аффинность IgG к АГ на 3-4 порядка выше, чем у IgM. Очень специфичный ответ именно на тот антиген, которым иммунизовали.
Клетки памяти
● Исходная численность каждого клона клеток памяти на 2–3 порядка выше, чем клона наивных клеток.
● Клетки памяти пребывают в клеточном цикле, а наивные лимфоциты — в фазе покоя (G0), для выхода из которой им требуется время и особые воздействия (активация).
● При действии АГ клетки памяти не должны проходить некоторые стадии развития (в случае В-клеток — переключение изотипов, мутагенез V-генов и созревание аффинитета; в случае Т-клеток — формирование связей TCR с корецепторами, перемещение TCR в рафты, дифференцировку Th1/Th2; для всех лимфоцитов — изменение спектра хемокиновых рецепторов).
● Клетки памяти быстрее и эффективнее реагируют на АГ (активируются, дифференцируются в эффекторные клетки), что обусловлено изменениями сигнальных путей. Так, изоформа CD45R0, характерная для клеток памяти, способствует более быстрой активации, чем CD45RA, свойственная наивным Т-клеткам. В результате Т-клетки памяти могут быть активированы при презентации антигена любыми АПК, тогда как наивные Т-клетки — только ДК.
● В силу более интенсивной рециркуляции и способности мигрировать в барьерные и нелимфоидные ткани Т-клетки памяти с большей вероятностью могут «встретить» АГ. Этот АГ может быть презентирован им на месте, а не только во вторичных лимфоидных органах, как в случае наивных Т-клеток.
6.14. Особенности физиологии и антигенных рецепторов B-1 и гамма-дельта-Т клеток, позволяющие отнести их к "врожденным лимфоцитам".
В1 клетки: IgM, IgA; нет соматического гипермутагенеза; синтез АТ спонтанный; специфичность АТ низкая; самоподдерживаются; сидят в серозных полостях и барьерных тканях; памяти нет (IgM всегда в одном кол-ве->каждый раз как первый)
ydTкл: сидят в барьерных тканях; для распознавания присутствие АГ в составе МНС необязательно
кто еще относится к «врожденным лимфоцитам»? – MZB, NKT, ILC.
6.15. Позитивная селекция CD4+ и CD8+ Т-клеток в тимусе. MHC-рестрикция.
См 6.11
- Сигнала с TCR нет -> +селекция в кортикальной зоне->CD4+/CD8+
- Слабый сигнал -> CD62L+CCR7+ наивные Ткл
- Средний сигнал -> FOXP3+CD4+CD25+ Tregs
- Сильный сигнал -> -селекция в медуллярной зоне
Развитие тимоцитов зависит от МНС
αβ Т-лимфоциты распознают фрагменты антигенов только в комплексе с молекулами MHC (MHC-рестрикция) (исключение - суперантигены);
МНС индивидуальны. Т-лимфоциты проходят позитивную селекцию на молекулах MHC, которые у каждого индивидуума свои, и пептидах, которые способны с этими аллелями связаться. У другого индивидуума набор молекул MHC будет другой, пептиды получатся тоже другие, в результате репертуар специфичностей зрелых лимфоцитов также будет отличаться. Абсолютная рестрикция: Т-клетки одной линии вообще "не видят" клеток-мишеней другой линии, несмотря на то, что они заражены тем же самым вирусом (МНС разные).
Смысл всего этого - что пептиды, узнаваемые в составе МНС для каждого будут разные, даже если исходник один.
Далее про процессинг АГ– обработка белка происходит через протеасому; потом МНС и пептиды экзоцитируются с помощью транспортных белков HLA-TAP, и инвариантная цепь заменяется на специфический пептид->представление Тх или Тк.
Дата: 2019-03-06, просмотров: 511.