У αβ Т-клеток есть α и β - две цепи, входящие в состав T-клеточного рецептора. Они распознают линейные пептидные эпитопы. Они находятся внутри белка, эти белки расщепляются, фрагмент белка в составе молекулы MHC (=HLA у человека) экспонируется на поверхности клетки. Такой комплекс чужого в контексте своего узнается Т-клеточным рецептором.
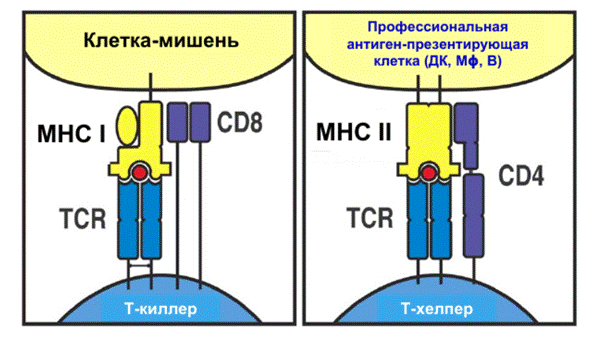
MHC бывают двух классов:
●MHCI (одна трансмембранная цепь + бета глобулин) встречается почти на всех клетках нашего организма, превращает их в мишень в случае, если в клетках появляются вирусные или опухолевые белки. MHC I распознается TCR, а о том, что это MHC I, Т-клетке сообщает корецептор CD8, сидящий на Т-киллере
●MHCII (две трансмембранные цепи) находится на поверхности антиген-презентирующих клеток, распознается TCR Т-хэлпера вместе с корецептором CD4
МНЕМОНИЧЕСКОЕ ПРАВИЛО:
MHCI - одна трансмембранная цепь
MHCII - две трансмембранные цепи
1х8 (MHCI и CD8)= 2х4 (MHCII и CD4)
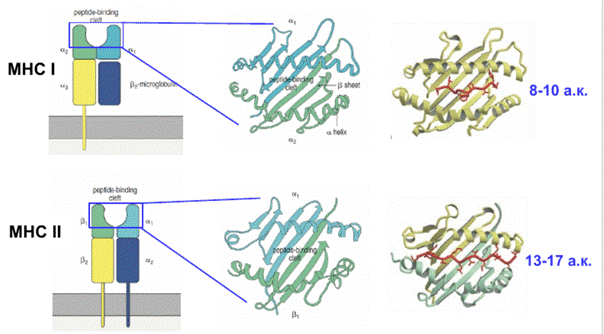
Есть различия в том, какие пептиды включаются в МНС. Это связано со структурой антигенсвязывающей бороздки Если посмотреть на антигенсвязывающую бороздку сверху, то можно увидеть, что это действительно борозда с дном из b-слоев и бортиками из a-спиралей. Бороздка MHCI более закрыта с обоих концов, в нее помещается пептид длиной до десяти аминокислот, а MHCII принимает в себя более длинные пептиды с большим разбросом по длине.
5.7. Презентация пептидов на молекулах MHC. Полиморфизм молекул MHC и его биологический смысл[33] .
МHCI презентация
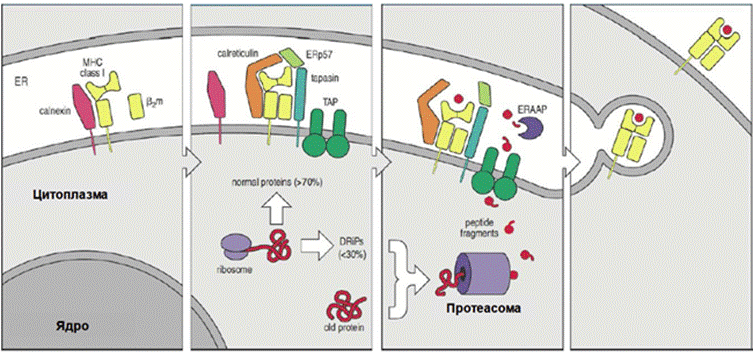
● Сборка молекулы MHCI из aльфа-цепи и бета2-микроглобулина осуществляется в ЭПС с участием специализированных белков-помощников - шаперонов.
● Предсобранная молекула MHCI связывается с белками TAP (transporters associated with antigen processing). Ненужные цитоплазматические белки и продукты неудачной трансляции (defective ribosomal products, DRiPs) направляются для деградации в протеасому.
● Все клеточные белки так или иначе нарезаются протеасомой, часть получившихся пептидов не утилизируется, а транспортируется в ЭПС при помощи белков-транспортеров TAP., где связывается в пептид-связывающей борозде свежесинтезированного MHCI, который затем направляется в мембрану путем экзоцитоза.
MHCII презентация
● В ЭПС специализированный белок (инвариантная цепь (Ii)) связывается с MHCII, блокируя связывание эндогенных пептидов. Отщепляется кислая эндосома с МНС внутри. 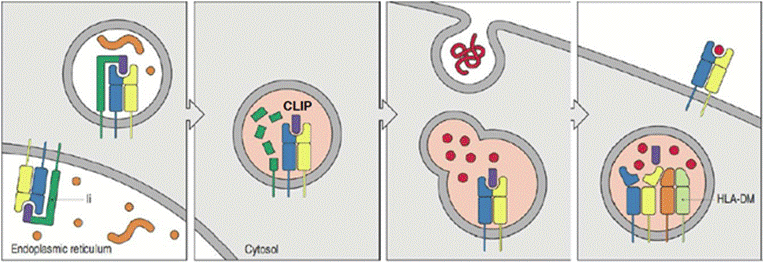
● В кислой эндосоме инвариантная цепь расщепляется, оставляя в комплексе с MHCII короткий пептид CLIP (class 2-associated invariant chain peptide).
● Антигены, попавшие в эндосому снаружи, деградируют до пептидов.
● Молекула HLA-DM связывается с MHCII, изменяя ее конформацию и освобождая от CLIP.
● Экзогенные пептиды связываются с MHCII и отправляются на поверхность путем экзоцитоза.
Кросс-презентация антигена
Антиген-презентирующие клетки умеют представлять антигенные пептиды в комплексе с молекулами MHCI, а не MHCII. Это явление называется кросс-презентацией, оно необходимо для эффективного цитотоксического ответа на вирусы, незаражающие антиген-презентирующие клетки, а также на злокачественные опухоли.
V(D)J рекомбинация. Какие локусы ей подвержены, какие генетические механизмы и какие ферменты задействованы. RSS, правило 12/23.
Физиологический смысл V(D)J перестройки генов иммуноглобулинов- это генерация разнообразия антиген-распознающих структур лимфоцитов, приводящий к формированию антиген-распознающих участков иммуноглобулинов и Т-клеточного рецептора. Гены иммуноглобулина и Т-клеточного рецептора состоят из повторяющихся сегментов, принадлежащих к трем классам: V (variable), D (diversity) и J (joining). В процессе V(D)J-перестройки генные сегменты, по одному из каждого класса, соединяются вместе.
● Белки RAG1 и RAG2 -эндонуклеазы, ответственные за включение процесса реаранжировки V-генов. Рекомбиназа RAG-1 димеризуются с RAG-2 в процессе перестройки V-генов и формирует RAG-эндонуклеазу, ответственную за осуществление двунитевых разрывов ДНК. Распознают и связываются с RSS (консервативные сигнальные последовательности, правило 23/12 в тяжелой цепи и 12/23 в легкой) и сближают их вместе. Затем инициируют двухцепочечный разрыв ДНК и делают шпильку - негомологичная рекомбинация.
Правило 12/23[34] : ориентация сигналов в генах тяжелых цепей иммуноглобулинов ,а также бета и гамма цепей TCR делает невозможной VJ рекомбинацию в этих локусах.
● Фермент TdT(терминальная дезоксинуклеотидилтрансфераза). обеспечивает нематричное добавление нуклеотидов при D-J рекомбинации - формирование N-вставок.
● Белки Ku70 и Ku86 - компоненты системы NHEJ – образующие гетеродимер, отвечают за удержание вместе фрагментов ДНК в зоне перестройки гена.
● ДНК-зависимая протеинкиназа и нуклеаза Artemis отвечают за «разрешение шпилек» (повторные однонитевые разрывы ДНК в участках ее Временного воссоединения)
● ДНК-лигаза IV отвечает за сшивку разрывов ДНК.
Дата: 2019-03-06, просмотров: 624.