Обозначение
Особенности
традиционное
ISBT
антигена
основа
750
Биохимия и генетика
Антигены LW представляют собой гликопротеины с мол. массой око-ло 40 кДа. Разрушение дисульфидных связей инактивирует вещество LW (Konigshaus, Holland [27]).
Ген LW картирован на коротком плече хромосомы 19 в позиции 19р13.3. Антигенный полиморфизм (LW a / LW b) обусловлен перемещением A 308 G в эк-зоне 1, что приводит к аминокислотной замене: глицин на аргинин в позиции 70 в первом IgSF-домене гликопротеина LW (Hermand и соавт. [21]).
Мол. масса гликопротеина, полученного посредством иммунопреципита-ции с использованием аллогенных анти-LW ab-антител (от миссис Big.), соответ-ствовала 37–47 кДа (Mallinson и соавт. [38], Bloy и соавт. [5, 6], Moore [39]). Субстрат, полученный при использовании моноклональных анти-LW ab-антител, имел меньшую мол. массу – от 36 до 43 кДа.
Мол. масса гликопротеина снижалась до 2 и 17 кДа после обработки N- и О-гликаназами соответственно (Bloy и соавт. [5]). Добавление Na2-ЭДТА (три-лон Б) к эритроцитам ингибировало антигены LW (Bloy и соавт. [6]). Ионы Mg2 + восстанавливали активность антигенов LW, ионы Mn2 + и Ca2 + были инертны.
Основываясь на результатах сравнительного исследования гликопротеина LW и протеина Rh с помощью химотриптического йодпептидного картирова-ния, Bloy и соавт. [5, 7] высказали предположение, что гликопротеин LW мо-жет являться гликозилированной формой Rh-протеина или, иными словами, Rh-полипептид является субстанцией-предшественником гликопротеина LW. Протеин Rh с мол. массой 31 кДа преципитировался одновременно с гликопро-теином LW, и это свидетельствовало, что указанные структуры эритроцитарной мембраны тесно связаны.
Bailly и соавт. [1] частично воспроизвели аминокислотную последователь-ность гликопротеина LW, что позволило создать олигонуклеотидные прайме-ры, исследовать кДНК и установить, что кодируемый пептид имеет мол. массу 26,5 кДа. Кроличьи антитела к синтетическому пептиду, состоящему из 15 ами-нокислот, реагировали в непрямой антиглобулиновой пробе со всеми образца-ми эритроцитов за исключением LW(a −b −). Эритроциты D + реагировали ин-тенсивнее, чем эритроциты D −. Эритроциты D + LW(a −b + ) давали слабовыра-женные реакции.
| MGSLFPLSLL FFLAAAYPGV GSALGRRTKR -01 | |
| AQSPKGSPLA PSGTSVPEWV RMSPEFVAVQ PGKSVQLNCS NSCPQPQNSS | 50 |
| LRTPLRQGKT LRGPGWVSYQ LLDVRAWSSL AHCLVTCAGK TRWATSRITA 100 | |
| YKPPHSVILE PPVLKGRKYT LRCHVTQVFP VGYLVVTLRH GSRVIYSESL 150 | |
| ERFTGLDLAN VTLTYEFAAG PRDFWQPVIC HARLNLDGLV VRNSSAPITL | 200 |
| MLAWSPAPTA LASGSIAALV GILLTVGAAY LCKCLAMKSQ A | 241 |
Рис. 18.1. Аминокислотная последовательность протеина LW.
Как показали Bailly и соавт. [1], Hermand и соавт. [21], ген LW кодирует протеин, включающий 271 аминокислоту (рис. 18.1), в том числе сигнальный
751
пептид (30 аминокислот), экстрацеллюлярный N-терминальный домен (208 аминокислот), трансмембранный гидрофобный домен (21 аминокислота) и С-терминальный цитоплазматический домен (12 аминокислот). Имеется четыре потенциальных участка N-гликозилирования: 38, 48, 160 и 191, которые зани-мает аспарагин. N-гликозилирование этих участков приводит к формированию гликопротеина с мол. массой 38–46 кДа.
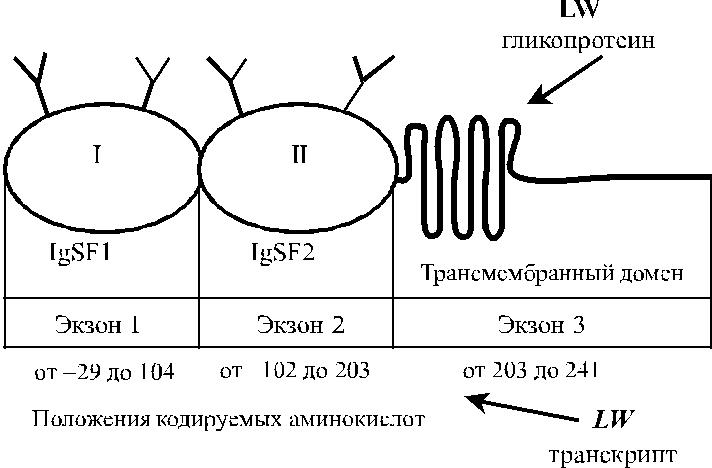
Рис. 18.2. Строение гликопротеина LW и транскрипта гена LW.
Гликопротеин LW включает два IgSF-домена (рис. 18.2) и структур-но связан с молекулами межклеточной адгезии ICAM-1, ICAM-2 и ICAM-3. Предложена трехмерная модель гликопротеина LW (Hermand и соавт. [22], Spring и соавт. [59]).
Ген LW имеет величину 2,65 кб, организован в виде трех экзонов (см. рис. 18.2) Экзон 1 кодирует нетранслируемую последовательность из 96 ко-донов, сигнальный пептид и первый IgSF-домен. Экзоны 1 и 2 отделены друг от друга интроном из 129 пар оснований. Еще один интрон (147 пар основа-ний) разделяет экзоны 2 и 3. Экзон 2 кодирует второй IgSF-домен. Экзон 3 ко-дирует трансмембранный и интрацеллюлярный домены и содержит в области 3ʹ нетранслируемую последовательность. Промоторная область гена содержит участки, влияющие на экспрессию антигенов LW на других клетках (Hermand и соавт. [23]).
Фенотипы LW a , LW b и LW(a −b −)
и большинстве популяций антигены LW a и LW b имеют противоположный характер распределения: LW a встречается часто, LW b – редко. Наибольшая ча-стота антигена LW b зарегистрирована у латышей и литовцев, в связи с чем этот фактор относят к своеобразным балтийским маркерам (табл. 18.3) и его обнару-жение в других популяциях, как полагают Sistonen и соавт. [55], отражает сте-пень влияния прибалтийских народов.
752
а финнов частота генов, генотипов и фенотипов соответствует следующим величинам:
| LW a | 0,971 | LW a / LW a | LW(a +b −) | 0,9429 |
| LW a / LW b | LW(a +b + ) | 0,0563 | ||
| LW b | 0,029 | LW b / LW b | LW(a −b + ) | 0,0008 |
До обнаружения антител анти-LW b практически все индивиды с редким фено-типом LW(a −) выявлялись в связи с присутствием в сыворотке их крови анти-LW-антител. Посемейные исследования часто не подтверждали наследственную пере-дачу гена LW (Race и Sanger [48], Beck [3], Giles [18]). Позднее было установлено, что LW a и LW b являются кодоминантными аллелями (Sistonen и соавт. [52, 54]) и не зависят от локуса RH (Levine и соавт. [35], Swanson и соавт.[60, 63], White и соавт. [67]). Обследование членов нескольких финских семей подтвердило это заключение.
Эксперименты по трансфекции кДНК LW a и LW b в клетки линии COS-7, выполненные Hermand и соавт. [21], показали, что моноклональные анти-LW ab-антитела более интенсивно реагируют с антигеном LW(a + ), чем с анти-геном LW(b + ).
Фенотип LW(a −b −) крайне редкий. DeVeber и соавт. [14] исследовали эритро-циты 10 552 канадцев с использованием сывороток анти-LW ab и не нашли ни одно-го с фенотипом LW(a −b −). Миссис Big., первая женщина, у которой были выявле-ны анти-LW ab-антитела, и ее брат имели фенотип LW(a −b −). Эритроциты всех 5 ее детей реагировали с ее сывороткой (DeVeber и соавт. [14], Sistonen и Tippett [54]). Как установили Hermand и соавт. [23], фенотип миссис Big. был обусловлен де-лецией кодонов 86–89 в экзоне 1 гена LW, что приводило к синтезу укороченного протеина, лишенного трансмембранного и цитоплазматического домена.
| Таблица 18.3 | |||
| Частота аллеля LW b у различных народов* | |||
| Популяция | Количество обследованных | Частота гена LW b | |
| Латыши | 677 | 0,059 | |
| Литовцы | 829 | 0,057 | |
| Эстонцы | 800 | 0,040 | |
| Финны | 6270 | 0,029 | |
| Русские (Вологодская область) | 383 | 0,022 | |
| Поляки (Польша и США) | 747 | 0,020 | |
| Шведы (Готланд) | 199 | 0,010 | |
| Шведы (Лунд) | 395 | 0,003 | |
| Венгры (Будапешт) | 421 | 0,004 | |
| Швейцарцы | 502 | 0,001 | |
| Бельгийцы (Льеж) | 211 | 0 | |
| Японцы (Осака) | 500 | 0 | |
| Африканцы (Сомали) | 1020 | 0 | |
*по Sistonen и соавт. [55].
753
Еще один человек с фенотипом LW(a −b −), житель Новой Гвинеи, выявлен Poole и соавт. [47]. В сыворотке его крови присутствовали анти-LW ab-антитела. Его сестра имела такой же фенотип – LW(a −b −); эритроциты сына были LW(a +b −) с нормально выраженными антигенами LW a и LW ab; у дочери указан-ные антигены были выражены слабо.
Экспрессия антигенов LW
Измерение участков связывания антигена LW с анти-LW ab-антителами пока-зало, что на одном эритроците взрослого человека D + присутствует 4400 LW ab-эпитопов, на одном эритроците человека D − находится 2835 LW ab-эпитопов. Для новорожденных D + и D − эти показатели составили 5150 и 3620 соответ-ственно (Mallinson и соавт. [38]).
Сообщалось, что антигены СсЕе, а также гомозиготность по гену D не ока-зывают какого-либо влияния на экспрессию антигенов LW (Swanson и соавт. [61]). Однако имеются и прямо противоположные данные. Так, Gibbs [17] на-шел, что эритроциты лиц DCe / DcE имели более сильные антигены D и LW, чем эритроциты лиц DcE / dce и DСе / dce. На эритроцитах лиц D u антигены D и LW были выражены еще более слабо (Swanson и соавт. [61]).
Антигены системы LW лучше выражены на эритроцитах новорожденных, поэ-тому реакция ксеногенных антител с эритроцитами детей более сильная, чем с эри-троцитами взрослых. В отличие от ксеногенных сывороток сыворотки анти-LW ал-логенного происхождения реже выявляют это различие (Swanson и соавт. [61]).
В онтогенезе антигены LW формируются на стадии колониеобразующих предшественников (Southcott и соавт. [58]) или позднее, на стадии проэритро-
бластов (Bony и соавт. [8], Hermand и соавт. [23]).
Антигены системы LW обладают некоторым эффектом дозы, который отме-тили Sistonen и соавт. [52], однако слабые реакции наблюдались с эритроцитами не только гетерозигот (LW а / LW b), но и гомозигот (LW а / LW а). При обследовании 10 014 финнов с фенотипом LW(а + ) эти авторы наблюдали слабовыраженные ре-акции у 722 человек, из которых 348 были гомо- и 374 гетерозиготами. Daniels
в полагает, что для экспрессии антигенов LW большее значение имеет выра-женность антигена D, чем гомо- или гетерозиготность по генам LW а и LW b.
Антигены системы LW устойчивы к действию папаина, фицина, трипсина и химотрипсина, однако разрушаются проназой (Lomas, Tippett [37]). Эту особен-ность используют для дифференцировки антител системы LW и Rh.
Экспрессия антигенов LW обусловлена не только генетическими факторами. Описан транзиторный фенотип LW − с временной утратой эритроцитами всех ан-тигенов LW. Этот фенотип диагностировался в связи с обнаружением антител к ан-тигенам LW a или LW ab. Утрата антигенов часто имела обратимый характер. Первое описание транзиторного фенотипа LW − у Rh-отрицательной беременной опубли-ковали Giles и Lundsgaard [19]. Незадолго до родов эритроциты беременной были LW −, а сыворотка крови содержала антитела анти-D, анти-С и анти-LW; прямая
754
антиглобулиновая проба с ее эритроцитами была слабоположительной. Через 1 год после родов эритроциты женщины тестировались как LW +, антитела анти-LW не выявлялись. Позднее было описано еще три случая транзиторного фенотипа LW −с анти-LW-антителами: у двух беременных D − и одного реципиента D + (Chown и со-авт. [11]). Авторы полагали, что имела место утрата антигенов LW, которую нельзя было объяснить блокадой LW-эпитопов антителами анти-LW.
11 из 18 мужчин с D −, иммунизированных антигеном D, Chown и соавт.
с наблюдали появление антител, напоминавших по специфичности анти-LW. Авторы предположили, что анти-LW-антитела, возможно, являются предвест-ником последующей выработки анти-D-антител.
Swanson и соавт. [62] наблюдали мальчика D +, которому была произведена транс-плантация костного мозга от родной сестры D −. Через 3 мес. после трансплантации в сыворотке крови реципиента определялись антитела анти-D и анти-LW. Вероятно, они образовались в результате иммунного ответа трансплантированных лимфоцитов донора на антигены D и LW реципиента. Спустя 2 года после трансплантации у реци-пиента выявлялись только слабые анти-D-антитела, анти-LW-антитела исчезли.
Экспрессия антигенов LW может снижаться при некоторых заболеваниях и возвращаться к норме по мере выздоровления. Данный феномен описан у не-скольких больных лимфомой, лейкемией, саркомой и другими злокачествен-
ными опухолями (Giles и соавт. [18], Perkins и соавт. [44], Villalba и соавт. [64], Komatsu и соавт. [26]). Нередко эти больные умирали до полного восстановле-ния экспрессии указанных антигенов на эритроцитах.
Komatsu и соавт. [26] описали японца со злокачественной лимфомой, у кото-рого во время двух рецидивов заболевания на фоне химиотерапии исчезал анти-ген LW а и появлялись анти-LW a-антитела. Во время ремиссий указанные анти-тела отсутствовали, эритроциты больного были LW(а + ).
Возникновение фенотипа LW − наблюдали во время беременности и имму-нодефицитного состояния (Reid и соавт. [49], Devenish и соавт. [15]).
Эритроциты большинства людей с приобретенным фенотипом LW(a −b −) не реагируют с антителами анти-LW ab.
Chown и соавт. [11] отметили связь между степенью экспрессии антигенов LW на эритроцитах и спектром антител в сыворотке крови. Некоторые больные LW(а −b −) содержали слабый антиген LW ab и анти-LW a-антитела. У других пациен-тов, утративших антигены LW, антитела имели направленность анти-LW ab. Нередко направленность антител (анти-LW a или анти-LW ab) установить не удавалось.
Sistonen и соавт. [52] описали больного LW(a −)LW ab +, у которого в терми-нальной фазе заболевания фенотип сменился на LW(a −)LW ab −. Два его брата и две дочери имели фенотип LW(a −)LW ab +. Больной был женат на двоюрод-ной сестре. Впоследствии некоторые члены указанной семьи были обследова-ны на наличие антигена LW b. Оказалось, что один из братьев больного и обе дочери имели фенотип LW(b + ). Эритроциты больного, хранившиеся длитель-ное время в замороженном состоянии, также были исследованы на наличие
755
антигена LW b. Адсорбционные тесты показали следы антигена, из чего сле-довал вывод, что антиген LW b мог быть утрачен вместе с другими антигенами этой системы.
Анти-LW-антитела
Сыворотки анти-LW a и анти-LW ab реагируют сильнее, если антигены LW a и LW ab находятся на эритроцитах D +. Те же антигены на эритроцитах D − реаги-руют слабее (Levine и Celano [32], Swanson и соавт. [60], DeVeber и соавт. [14], White и соавт. [67]). В некоторых случаях эти различия настолько выражены, что антитела анти-LW a ошибочно идентифицируют как анти-D.
Анти-LW b-антитела проявляют себя в серологических реакциях с эри-троцитами D + и D − так же, однако из-за низкой частоты открываемого ими LW b-антигена их трудно принять за анти-D.
Аллоиммунные анти-LW a-антитела обнаруживают у лиц, имеющих фенотип LW(а −b + ) и LW(а −b −).
Большинство анти-LW a-антител найдены у пациентов, которым производили гемотрансфузии, однако сенсибилизация может наступить вследствие беремен-ности (Giles [18]).
Napier и Rowe [41] находили анти-LW a-антитела у добровольцев D −, искус-ственно иммунизированных антигеном D.
Известно всего два случая образования анти-LW ab-антител у лиц с генети-чески обусловленным (унаследованным от родителей) фенотипом LW(а −b −) (DeVeber и соавт. [14], Poole и соавт. [47]). В одном из них упомянутая выше женщина (миссис Big.) имела три беременности и не подвергалась гемотранс-фузиям. Сыворотка ее крови содержала сильные анти-LW ab-антитела, которые реагировали с эритроцитами LW ab + D + в разведении 1 : 32 000 и с эритроцита-ми LW ab + D − в разведении 1 : 1000. При последующем наблюдении титр анти-тел снизился, они перестали улавливать различия в экспрессии антигена LW ab на эритроцитах D + и D − (DeVeber и соавт. [14]).
Perrault и соавт. [45] отделили анти-LW ab-антитела IgM от анти-LW ab-антител IgG и установили, что агглютинины IgM лучше выявляют различия в экспрес-сии антигена LW ab на эритроцитах D + и D −, чем антитела IgG.
Несколько образцов анти-LW b-антител было найдено в Финляндии. Первый из них содержал моноспецифические анти-LW b-антитела, последующие при-сутствовали в сочетании с антителами анти-K, анти-Kр а и анти-Ul a. Все финны, имевшие анти-LW b-антитела, были D +, у них наблюдались заболевания крови, и им многократно производили гемотрансфузии.
Антитела системы LW могут иметь аутоиммунное происхождение. Их об-наруживали у больных LW +, страдавших аутоиммунной гемолитической ане-мией. Эритроциты пациентов реагировали в прямой антиглобулиновой пробе, что свидетельствовало о присутствии на их поверхности аутоантител. Свободно циркулирующие аутоантитела, имевшиеся у больных, проявляли себя как
756
аллоиммунные, и их трудно было отличить от истинных аллоиммунных, осо-бенно у больных с приобретенным фенотипом LW −.
Случаев развития ГБН или посттрансфузионных реакций, обусловленных LW-антителами, не зарегистрировано. Многим реципиентам, имевшим антите-ла анти-LW a и анти-LW ab, производили трансфузии эритроцитов, несовместимых по системе LW, без каких-либо проявлений (Perkins и соавт. [44], Komatsu и со-авт. [26], Reid и соавт. [49], Devenish [15], Cummings и соавт. [12], Chaplin и соавт. [10]). Суперактивные анти-LW ab-антитела, обнаруженные у миссис Big., не оказа-ли влияния на здоровье родившегося у нее ребенка (DeVeber и соавт. [14]).
Антитела системы LW чаще относятся к IgG, а именно к IgG1-субклассу (Reid и соавт. [49], Napier и Rowe [41], Cummings и соавт. [12]). Один из описан-ных образцов анти-LW ab-антител представлял собой смесь IgМ и IgG, другой, не активный в антиглобулиновой пробе, содержал антитела только IgМ-класса (Swanson и соавт. [63]).
Приживаемость несовместимых по LW-антигенам эритроцитов у большин-ства реципиентов, имевших анти-LW a- и анти-LW ab-антитела, существенно не отличалась от контрольных показателей (Komatsu и соавт. [26], Reid и со-авт. [49], Napier и Rowe [41], Cummings и соавт. [12], Chaplin и соавт. [10]). Исключением явились два образца анти-LW ab-антител, которые относились к IgG3-субклассу (Villalba и соавт. [64], Herron и соавт. [24]). В одном случае при-живаемость эритроцитов LW + D −, меченных радиоактивными изотопами, со-ставила 53 % уже через 1 ч после их введения.
с реципиентов, содержащих высокоактивные анти-LW b-антитела, эритроци-ты LW(b + ), введенные внутривенно, быстро исчезали из кровотока: полупери-од их циркуляции составил от 2 до 5 ч (Sistonen и соавт. [53]).
По результатам экспериментов некоторые образцы антител LW могли быть отнесены к трансфузионно опасным, однако видимых реакций они не давали.
Perrault [45] при обследовании 45 000 доноров выявил 10 человек, имев-ших аутоантитела анти-LW. Восемь носителей аутоантител были здоровы, 2 больны (у одного был язвенный колит, у другого – опухоль слюнной железы). Аутоантитела выявлены с помощью автоматического анализатора в услови-ях низкоионной среды с полибреном, при использоваии обычных методов ис-следования их обнаружить не удалось. Уменьшения продолжительности жизни эритроцитов in vivo аутоантитела не вызывали.
Levine [29] высказал суждение, что аутоантитела анти-LW у больных c ау-тоиммунной гемолитической анемией, у которых получены положительные результаты прямой пробы Кумбса, встречаются весьма часто. Celano и Levine [9], адсорбируя элюаты, снятые с эритроцитов 6 таких больных, эритроцитами LW(a −b + ), нашли аутоантитела анти-LW a во всех случаях, один элюат содер-жал только LW a-аутоантитела.
Другие авторы (Vos и соавт. [65]) для адсорбции элюатов использовали эри-троциты LW(a −b −)LW ab − и выявили аутоантитела анти-LW у 6 из 8 больных
757
c аутоиммунной гемолитической анемией. Во всех элюатах аутоантитела анти-LW присутствовали с антителами другой специфичности.
Антитела системы LW были получены путем иммунизации кроликов и, гораздо успешнее, морских свинок. Для иммунизации использовали эри-троциты обезьян Macacus rhesus, бабуинов, а также людей с различной Rh-принадлежностью (Fisk и Foord [16], Levine и соавт. [32, 36], Swanson и соавт. [61], Polesky и соавт. [46], Wiener и соавт. [68, 69]). Экстракты, полученные из эритроцитов человека путем подогрева, также стимулировали у животных обра-зование анти-LW-антител (Murray и Clark [40], Levine и соавт. [30, 31]).
Антитела, вырабатываемые морскими свинками и кроликами, не реагируют с эри-троцитами LW(a −b + ), проявляя таким образом специфичность анти-LW a. Данные о способности указанных животных вырабатывать анти-LW ab-антитела отсутствуют.
Эритроциты LW(a −b + ) и LW(a −b −) от миссис Big. стимулировали образова-ние анти-LW-антител у морских свинок (Vos и соавт. [65], Polesky и соавт. [46]). По отношению к этим животным лишь эритроциты Rhnull не обладали иммуно-генностью (Levine и соавт. [29, 34], Polesky и соавт. [46]). Иммунный ответ живот-ных на введение эритроцитов LW(a −b −) от мисс Big. оказался полной неожидан-ностью, поскольку на указанных клетках LW-гликопротеины отсутствовали.
в настоящему времени имеется несколько серий моноклональных анти-LW ab-антител, которые были получены путем иммунизации мышей эритроци-тами человека и обезьян Macacus rhesus (Sonneborn и соавт. [56, 57], Oliveira и соавт. [42]). Антитела взаимодействуют со всеми образцами эритроцитов за ис-ключением LW(a −b −). Некоторые образцы антител реагировали с папинизиро-
ванными эритроцитами LW(a −b + ) (Sonneborn и соавт. [56]).
Связывание мышиных моноклональных анти-LW ab-антител полностью бло-кировалось после контакта эритроцитов с аллогенными антителами указанной специфичности. Частичный блокирующий эффект давали также человеческие анти-LW a-сыворотки. Сыворотки анти-D реакцию анти-LW ab-антител не блоки-ровали, что свидетельствовало о разных качествах антигенов LW и RH.
Hermand и соавт. [22] полагают, что мышиные МКА распознают эпитопы, на-ходящиеся на первом IgSF-домене LW-гликопротеина. Получены также мыши-ные МКА, распознающие антигенные участки на первом и втором IgSF-доменах (Blanchard и соавт. [4]). Иммунизацию мышей проводили рекомбинантным химер-ным протеином, состоящим из двух IgSF-доменов и Fc-фрагмента IgG1 человека.
Дата: 2019-02-24, просмотров: 420.