| Фенотип | Замена нуклеотидов | Экзон | Замена аминокислот |
| Do a / Do b | A 793 G | 2 | Asn 265 Asp |
| Hy +/Hy − | G 323 T | 2 | Gly 108 Val |
| Jo(a + )/Jo(a −) | C 350 T | 2 | Thr 117 Ile |
РНК-транскрипты гена DO обнаруживали в селезенке, лимфатических узлах, костном мозге и эмбриональной печени ; в тимусе и лейкоцитах пери-ферической крови они отсутствовали (Gubin и соавт. [9]). Они не выявлялись
51. течение первых 4 дней культивирования клеток периферической крови чело-века в присутствии эритропоэтина (Gubin и соавт. [9]).
Антитела системы Dombrock
После открытия антигенов Do a и Do b [23, 45] последовала серия сообщений
82. выявлении новых образцов антител анти-Do a (Webb и соавт. [53], Williams, Crawford [54], Polesky и соавт. [31], Moulds и соавт. [25], Kruskall и соавт. [17], Judd, Steiner [15], Roxby и соавт. [39], Strupp и соавт. [44]) и анти-Do b (Strupp и соавт. [44], Yvart и соавт. [55], Moheng и соавт. [22], Halverson и соавт. [11], Shirey и соавт. [41]).
Частота анти-Do a-антител в исследованных выборках практически не отли-чалась от частоты антител анти-Do b и соответствовала ожидаемой, рассчитан-ной по частоте генов. На основании этого исследователи пришли к выводу, что антигены Do a и Do b одинаковы по своей способности вызывать аллоиммуни-зацию (Issitt, Anstee [13]). Антитела Dombrock чаще сопутствовали антителам другой специфичности, однако известны также сыворотки, содержащие моно-специфические анти-Do a-антитела (Moulds и соавт. [25], Roxby и соавт. [39]) и моноспецифические анти-Do b-антитела (Shirey и соавт. [41]). О выявлении естественных антител Dombrock не сообщалось.
Polesky и соавт. [31] наблюдали женщину, у которой анти-Do a-антитела обра-зовались во время первой беременности. Ребенок имел фенотип Do(a + ).
Антитела анти-Do a и анти-Do b относятся к классу IgG, не обладают способ-
ностью связывать комплемент (Polesky, Swanson [30], Moulds и соавт. [25], Yvart
и соавт. [55]). Их ни разу не описывали как причину ГБН. В отдельных случаях эти антитела обусловливали положительную прямую антиглобулиновую пробу с эритроцитами новорожденных без гемолитической реакции (Polesky и соавт. [31], Moulds и соавт. [25], Yvart и соавт. [55]).
Вместе с тем анти-Do a-антитела вызывали как острые, так и замедленные посттрансфузионные реакции, особенно у больных с серповидно-клеточной анемией (Kruskall и соавт. [17], Judd, Steiner [15], Strupp и соавт. [44]). Такие же реакции вызывали и анти-Do b-антитела (Strupp и соавт. [44], Moheng и соавт. [22], Halverson и соавт. [11], Shirey и соавт. [41]).
734
Тесты на приживление эритроцитов in vivo, а также эксперименты с моно-слоем моноцитов in vitro, показали, что антитела Dombrock ускоряют разруше-ние эритроцитов, несущих антигены Dombrock (Polesky, Swanson [30], Shirey и соавт. [41]).
// одном случае, описанном Gudino и соавт. [10], у пациента, имевше-го анти-Do a-антитела, приживаемость эритроцитов Do(a + ) была нормальной. Переливание ему Do a-положительных эритроцитов не вызвало реакции.
// некоторых случаях антитела анти-Do a и анти-Do b выявляли post factum, когда трансфузии уже были выполнены. В отдельных случаях эти антитела трактовали как слабые аллогенные, не имеющие клинического значения, ауто-
иммунные или HLA-антитела (Kruskall и соавт. [17], Judd, Steiner [15], Strupp и соавт. [44], Halverson и соавт. [11]).
// противоположность антигенам Do a и Do b антиген Gy a более иммуногенен
110. выраженно проявляет себя при разногруппной беременности женщин Gy(a −) (Swanson и соавт. [46], Race, Sanger [32], Clark и соавт. [6], Okubo и соавт. [29]).
Антитела анти-Gy a, анти-Ну и анти-Jo a естественного происхождения не опи-саны (Daniels [7]). Однако могут быть исключения. Ellisor и соавт. [8] выявили анти-Gy a-антитела у пожилого мужчины, которому не проводили гемотрансфу-зий. Через 3 мес. антитела исчезли. Эритроциты этого человека, фенотип которо-го был определен как Gy(a −), адсорбировали анти-Ну-антитела, которые можно было затем элюировать. Стандартные эритроциты Gy(a −) такой способностью не обладали. Это дает основания полагать, что фенотип Gy(a −) может быть приобре-тенным (Reid и соавт. [34]). Процесс подобной трансформации, по-видимому, мо-жет сопровождаться появлением соответствующих транзиторных антител.
Антитела анти-Gy a, анти-Ну и анти-Jo a, как правило, относятся к классу IgG (Swanson и соавт. [46], Clark и соавт. [6], Moulds и соавт. [26], Beattie, Castillo [3], Hsu и соавт. [12], Jensen и соавт. [14], Morel и соавт. [24], Brown и соавт. [4], Ellisor и соавт. [8], Barrett и соавт. [2]). Один образец анти-Gy a-антител содер-жал фракцию IgA и, как удалось установить с помощью антиглобулиновой про-бы, связывал комплемент (Clark и соавт. [6]). Один образец анти-Ну-антител, вызывавший прямую агглютинацию эритроцитов, помимо IgG, содержал фрак-
цию IgМ (Barrett и соавт. [2]).
Beattie и Castillo [3] описали случай гемолитической посттрансфузионной реакции у мужчины, имевшего анти-Ну-антитела, которому были перелиты две дозы эритроцитов Ну +.
Описан также реципиент с наличием анти-Gy a-антител, которому были про-изведены трансфузии 10 доз эритроцитов Gy(a + ) (Mak и соавт. [20]). Каких-либо проявлений несовместимости при этом авторы не наблюдали.
№ больного с транзиторными анти-Gy a-антителами приживаемость эритро-цитов Gy(a + ) in vivo была нормальной (Hsu и соавт. [12]). В аналогичных ис-следованиях ускоренную элиминацию эритроцитов in vivo вызывали антитела анти-Ну (Hsu и соавт. [12]) и анти-Jo a (Viggiano и соавт. [50]).
735
100. последние годы Rao и соавт. [33] получили мышиные МКА, реагирующие со всеми антигенами Dombrock, но дающие слабые реакции с эритроцитами Gy(a −).
Биологическая функция
Продукт аллеля Do b содержит аминокислотную последовательность Arg – Gly – Asp, характерную для молекул клеточной адгезии. Аллель Do a кодирует
125. том же участке фрагмент с другой последовательностью аминокислот, Arg – Gly – Asn, что может повлиять на адгезивную способность субстрата.
Экзон 2 локуса DO содержит участок, характерный для генов, контролирующих синтез аденозиндифосфатрибозилтрансферазы (АДФ-трансферазы) (Koch-Nolte, Haag [16]). Возможно, гены Dombrock способны модифицировать этот фермент и таким образом влиять на его функциональную активность (Gubin и соавт. [9]).
Spring и соавт. [42, 43], Telen и соавт. [47] наблюдали у больных пароксиз-мальной холодовой гемоглобинурией две популяции эритроцитов, одна из кото-рых была лишена ГФИ-ассоциированных протеинов и не содержала антигенов Dombrock. Эти эритроциты были более чувствительны к комплементу.
Список литературы
157. Banks J.A., Hemming N., Poole J. Evidence that Gy a, Hy and Jo a antigens belong to the Dombrock blood group system // Vox Sang. – 1995. – V. 68. – P. 177–182.
158. Barrett V.J., O’Brien M.M., Moulds J.J. et al. Anti-Holley detected in a primary immune response // Immunohematology. – 1996. – V. 12. – P. 62–65.
159. Beattie K.M., Castillo S. A case report of a hemolytic transfusion reaction caused be anti-Holley // Transfusion. – 1975. – V. 15. – P. 476–480.
160. Brown D. Reactivity of anti-Jo a with Hy- red cells [Abstract] // Transfusion. – 1985. –
// 25. – P. 462.
161. Chandanayingyong D., Sasaki T.T., Greenwalt T.J. Blood groups in Thais // Transfusion. – 1967. – V. 7. – P. 269–276.
162. Clark M.J., Poole J., Barnes R.M. et al. Study of the Gregory blood group in an English family // Vox Sang. – 1975. – V. 29. – P. 301–305.
163. Daniels G.L. Human Blood Groups. – 2-nd ed. – Oxford: Blackwell Science, 2002. – 560 p.
164. Ellisor S.S., Reid M.E., Avoy D.R. et al. Transient anti-Gy a in an untransfused man: serologic characteristics and cell survival study // Transfusion. – 1982. – V. 22. – P. 166–168.
165. Gubin A.N., Njoroge J.M., Woida U. et al. Identification of the Dombrock blood group glycoprotein as a polymorphic member of the ADP-ribosyltransferase gene family // Blood. – 2000. – V. 96. – P. 2621–2627.
166. Gudino M., Kranwinkel N., Lenart S., Harrison L. Successful transfusion of Dombrock ([Do(a + )] red blood cells to a patient with anti-Do(a) [Abstract] // Transfusion. – 1986. –
// 26. – P. 546.
167. Halverson G., Shanahan E., Santiago I. et al. The first reported case of anti-Do b. causing an acute hemolytic transfusion reaction // Vox Sang. – 1994. – V. 66. – P.206–209.
168. Hsu T.C.S., Jagathambal K., Sabo B.H., Sawitsky A. Anti-Holley (Hy): characterization of another example // Transfusion. – 1975. – V. 15. – P. 604–607.
169. Issitt P.D., Anstee D.J. Applied Blood Group Serology. – 4-th ed. – Durham, NC, USA: Montgomery Sc. Publ., 1998. – 1208 p.
170. Jensen L., Scott E.P., Marsh W.L. et al. Anti-Jo a: an antibody defining a high-frequency erythrocyte antigen // Transfusion. – 1972. – V. 12. – P. 322–324.
736
158. Judd W.J., Steiner E.A. Multiple hemolytic transfusion reactions caused by anti-Do a // Transfusion. – 1991. – V. 31. – P. 477–478.
159. Koch-Nolte F., Haag F. Mono (ADP-ribosyl) transferases and related enzymes in animal tissues: emerging gene families // Adv. Exp. Clin. Biol. – 1997. – V. 419. – P. 1–13.
160. Kruskall M.S., Greene M.J., Strychrz D.M. et al. Acute hemolytic reaction due to anti-Dombrock (Do a) [Abstract] // Transfusion. – 1986. – V. 26. – P. 554.
161. Laird-Fryer B., Moulds M.K., Moulds J.J. et al. Subdivision of the Gy a –Hy phenotype [Abstract] // Transfusion. – 1981. – V. 21. – P. 633.
162. Lewis M., Kaita H., Giblett E.R., Anderson J.E. Genetic linkage analysis of the Dombrock (Do) blood group locus // Cytogenet. Cell Genet. – 1978. – V. 22. – P. 313–318.
163. Mak K.H., Lin C.K., Ford D.S. et al. The first example of anti-Gy a detected in Hong Kong // Immunohematology. – 1995. – V. 11. – P. 20–21.
164. Massaquoi J.M. Two further examples of anti-Gy a // Transfusion. – 1975. – V. 15. – P. 150–151.
165. Moheng M.C., McCarthy P., Pierse S.R. Anti-Do b implicated as the cause of a delayed hemolytic transfusion reaction // Transfusion. – 1985. – V. 25. – P. 44–46.
166. Molthan L., Crawford M.N., Tippett P. Enlargement of the Dombrock blood group system: the finding of anti-Do b // Vox Sang. – 1973. – V. 24. – P. 382–384.
167. Morel P., Myers M., Marsh W.L., Bergren M. The third example of anti-Jo a: inheritance of the Jo a red cell antigen [Abstract] // Transfusion. – 1976. – V. 16. – P. 531.
168. Moulds J., Futrell E., Fortez P., McDonald C. Anti-Do a: further clinical and serological observations [Abstract] // Transfusion. – 1978. – V. 18. – P. 375.
169. Moulds J.J., Polesky H.F., Reid M., Ellisor S.S. Observation on the Gy a and Hy antigens and the antibodies that define them // Transfusion. – 1975. – V. 15. – P. 270–274.
170. Nakajima H., Moulds J.J. Do a (Dombrock) blood group antigen in the Japanese: tests on further population and family samples // Vox Sang. – 1980. – V. 38. – P. 294–296.
171. Nakajima H., Skradski K., Moulds J.J. Do a (Dombrock) blood group antigen in Japanese // Vox Sang. – 1979. – V. 36. – P. 103–104.
172. Okubo Y., Nagao N., Tomita T. et al. The first examples of the Gy(a −) Hy– phenotype and anti-Gy a found in Japan // Transfusion. – 1986. – V. 26. – P. 214–215.
173. Polesky H.F., Swanson J. Studies on the distribution of the blood group antigen Do a (Dombrock) and the characteristics of anti-Do a // Transfusion. – 1966. – V. 6. – P. 268–270.
174. Polesky H.F., Swanson J., Smith R. Anti-Do a stimulated by pregnancy // Vox Sang. – 1968. – V. 14. – P. 465–466.
175. Race R.R., Sanger R. Blood Groups in Man. – 6-th ed. – Oxford: BSP, 1975. – 659 p.
176. Rao N., Udani M., Nelson J. et al. Investigations using a novel monoclonal antibody to the glycosylphosphatidylinositol-anchored protein that carries Gregory, Holley, and Dombrock blood group antigens // Transfusion. – 1995. – V. 35. – P. 459–464.
177. Reid M.E., Ellisor S.S., Sabo B. Absorption and elution anti-Hy from one of four Gy(a −) human red blood cell samples // Transfusion. – 1982. – V. 22. – P. 528–529.
178. Reid M.E., Lomas-Francis C. The Blood Group Antigen: FactsBook. – 2-nd ed. – London: Academic Press, 2004. – 561 p.
179. Rios M., Hue-Roye K., Lee A.H. et al. DNA analysis for Dombrock polymorphism // Transfusion. – 2001. – V. 41. – P. 1143–1146.
180. Rios M., Hue-Roye K., Miller J.L. et al. Molecular basis associated with the Dombrock null phenotype [Abstract] // Blood. – 2000. – V. 96. – P. 452a.
181. Rios M., Hue-Roye K., Oyen R., Reid M. Molecular basis of the Gy(a + w) Hy– phenotype [Abstract] // Transfus. Clin. Biol. – 2001. – V. 8 (Suppl.). – 14S.
182. Roxby D.J., Paris J.M., Stern D.A., Young S.G. Pure anti-Do a stimulated by pregnancy // Vox Sang. – 1994. – V. 66. – P. 49–50.
737
191. Schmidt R.P., Frank S., Baugh M. New antibodies to high incidence antigenic determinants (anti-So, anti-El, anti-Hy and anti-Dp) [Abstract] // Transfusion. – 1967. – V. 7. – P. 386.
192. Shirey R.S., Boyd J.S., King K.E. et al. Assessment of the clinical significance of anti-Do b // Transfusion. – 1998. – V. 38. – P. 1026–1029.
193. Spring F.A., Reid M.E. Evidence that the human blood group antigens Gy a and Hy are carried on a novel glycosylphosphatidylinositol-linked erythrocyte membrane glycoprotein
J. Vox Sang. – 1991. – V. 60. – P. 53–59.
194. Spring F.A., Reid M.E., Nicholson G. Evidence for expression of the Jo a blood group antigen on the Gya / Hy-active glycoprotein // Vox Sang. – 1994. – V. 66. – P. 72–77.
195. Strupp A., Cash K., Uelinger J. Difficulties in identifying antibodies in the Dombrock blood group system in multiply alloimmunized patients // Transfusion. – 1998. – V. 38. – P. 1022– 1025.
196. Swanson J., Polesky H.F., Tippett P., Sanger R. A ’new’ blood group antigen, Do a // Nature. – 1965. – V. 206. – P. 313.
197. Swanson J., Zweber M., Polesky H.F. A new public antigenic determinant Gy a (Gregory) // Transfusion. – 1967. – V. 7. – P. 303–307.
198. Telen M.J., Rosse W.F., Parker C.J. et al. Evidence that several high-frequency human blood group antigens reside on phosphtidylinositol-linked erythrocyte membrane protein // Blood. – 1990. – V. 75. – P. 1404–1407.
199. Tippett P. Genetics of the Dombrock blood group system // J. Med. Genet. – 1967. – V. 4. –
7–11.
200. Tippett P., Sanger R., Swanson J., Polesky H.F. The Dombrock blood group system // Proc. 10-th Congr. Europ. Soc. Haematol. – 1965. – V. II. – P. 382–384.
201. Viggiano E., Jacobson G., Zurbito F. A Chromium51 survival study on a patient with anti-‘Jo a / Jc a’ [Abstract] // Transfusion. – 1985. – V. 25. – P. 446.
202. Weaver T., Kavitsky D., Carty L. et al. An association between the Jo a and Hy phenotypes [Abstract] // Transfusion. – 1984. – V. 24. – P. 246.
203. Weaver T., Lacey P., Carty L. et al. Evidence that Jo a and Jc a are synonymous // Transfusion. – 1986. – V. 26. – P. 561.
204. Webb A.J., Lockyer J.W., Tovey G.H. The second example of anti-Do a // Vox Sang. – 1966. –
11. – P. 637–639.
179. Williams C.H., Crawford M.N. The third example of anti-Do a // Transfusion. – 1966. –
6. – P. 310.
180. Yvart J., Cartron J., Fouillade M.T. et al. Un nouvel exemple d’anti-Do b // Rev. Franc. Transfus. Immunohemat. – 1977. – V. 20. – P. 395–400.
738
Глава 17.
Система Colton
Система Colton (Колтон) представлена тремя антигенами. Два из них, Co a (частый) и Co b (редкий), находятся в антитетичных отношениях. Третий анти-ген, Со3, присутствует на всех эритроцитах, за исключением эритроцитов лиц
194. нулевым фенотипом, Co(a −b −) (табл. 17.1). Групповые детерминанты Colton расположены на белке, получившем название аквапорин-1 (AQP-1).
Генный локус СО картирован на коротком плече хромосомы 7. Нулевой фенотип Co(a −b −) может быть обусловлен моносомией по хромосоме 7. Антигенные различия Colton обусловлены одиночными мутациями гена СО, приводящими к замене аланина в позиции 45 валином.
Таблица 17.1
Антигены Colton
| Обозначение | Частота | Примечание | ||
| традиционное | ISBT | |||
| Co a | CO1 | Высокая | Антитетичен Co b, Ala 45 | |
| Co b | CO2 | Низкая | Антитетичен Co a, Val 45 | |
| Co3 | CO3 | Очень высокая | Отсутствует у лиц Co(a −b −) | |
Негликозилированный вариант AQP-1 имеет мол. массу 28 кДа, гликозили-рованный – от 40 до 60 кДа (Denker et al [14]. Количество молекул AQP-1 на одну клетку составляет 120–160 тыс.
Методом ПЦР с использованием олигонуклеотидных праймеров исследова-на кДНК клеток эмбриональной печени человека. Это позволило сначала уста-новить аминокислотную последовательность N-терминального участка аквапо-рина-1, а затем изолировать кДНК аквапорина-1 из библиотеки генов костно-мозговых клеток человека. Найден участок ДНК величиной 807 пар нуклео-тидов, кодирующий синтез полипептида из шести трансмембранных доменов (рис. 17.1 и 17.2). N- и С-терминальные участки полипептида расположены вну-три клетки (Preston, Agre [43]). Полипептид, несущий антигены Colton, состоит из 269 аминокислот (рис. 17.3).
Обе половины молекулы аквапорина (по три трансмембранных домена в каждой) имеют участки NPA со сходной последовательностью аминокислот. Согласно предложенной трехмерной модели, участки NPA между трансмем-бранными доменами образуют канал транспорта воды и функционируют как
739
единое целое (см. рис. 17.2) (Jung и соавт. [24], Murata и соавт. [37], De Groot
15 соавт. [12]).
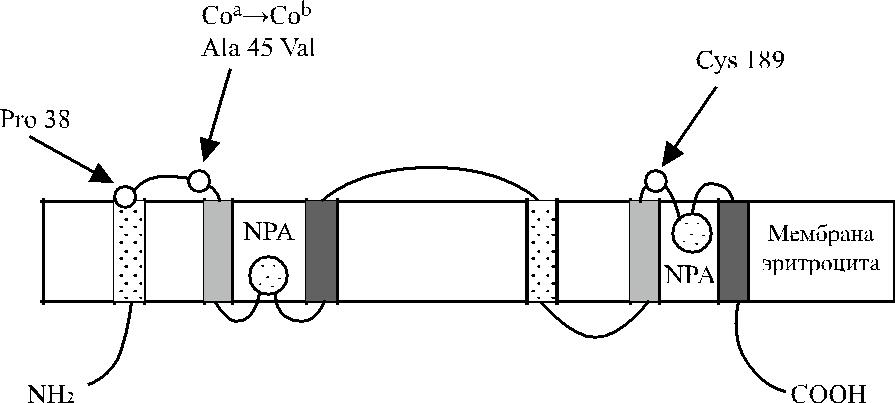
Рис. 17.1. Размещение антигенов Colton на мембране эритроцита.
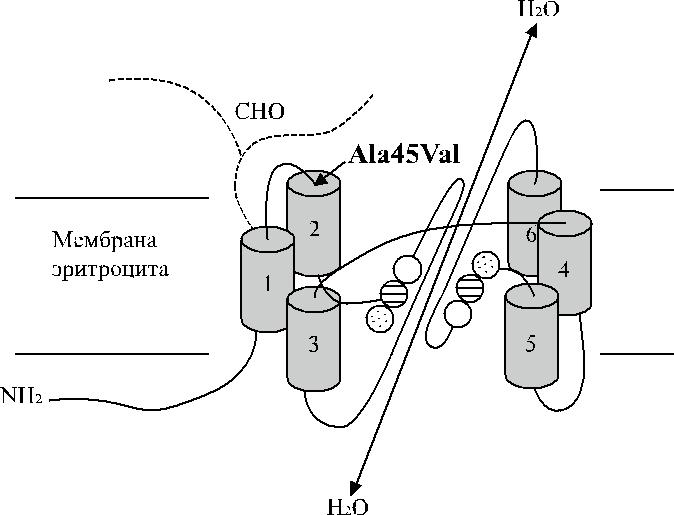
Рис. 17.2. Трехмерная модель аквапорина. Кружка-ми обозначены участки NPA.
| MASEFKKKLF WRAVVAEFLA TTLFVFISIG SALGFKYPVG NNQTAVQDNV | 50 |
| KVSLAFGLSI ATLAQSVGHI SGAHLNPAVT LGLLLSCQIS IFRALMYIIA | 100 |
| QCVGAIVATA ILSGITSSLT GNSLGRNDLA DGVNSGQGLG IEIIGTLQLV | 150 |
| LCVLATTDRR RRDLGGSAPL AIGLSVALGH LLAIDYTGGG INPARSFGSA | 200 |
| VITHNFSNHW IFWVGPFIGG ALAVLIYDFI LAPRSSDLTD RVKVWTSGQV | 250 |
| EEYDLDADDI NSRVEMKPK | 269 |
Рис. 17.3. Аминокислотная последовательность молекулы, несущей антигены Colton.
Первая экстрацеллюлярная петля молекулы аквапорина может быть N-гликозилирована и, подобно протеину полосы 3, обладать АВН-антигенной активностью (Smith и соавт. [52]).
Ген AQP-1 локализован в позиции 7р14, имеет величину 17 кб и представлен четырьмя экзонами (рис. 17.4), кодирующими аминокислоты в позициях 1–128, 129–183, 184–210 и 211–269 (Moon и соавт. [34], Umenishi, Verkman [57]).
740
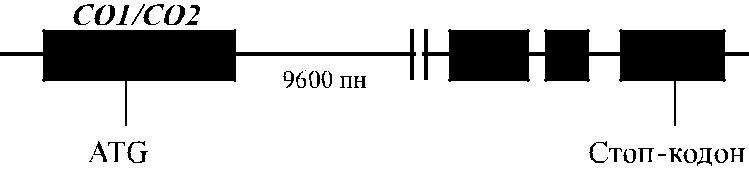
Рис. 17.4. Генная карта локуса CO (AQP-1).
Вскоре после обнаружения того факта, что локусы CO и AQP-1 находят-ся в одном и том же участке хромосомы 7, были получены новые данные. Установлено, что антитела анти-Со а и анти-Co b могут избирательно преципи-тировать молекулы аквапорина на эритроцитах, содержащих антиген Со а или Co b соответственно (Smith и соавт. [52]). Анти-Со3-антитела позволяли посред-ством иммунопреципитации выделить аквапорин из эритроцитов Co(a +b −) и Co(a −b + ).
Co a и Co b
C 1967 г. Heisto и соавт. [19] сообщили о выявлении трех образцов сыворо-ток, открывавших новый антиген с высокой частотой встречаемости. Антиген был обозначен как Co a, или Colton, по имени первого пробанда. Любопытен тот факт, что в действительности его фамилия была Calton, но она, как отметили Reid и Lomas-Francis [46], была неразборчиво написана на пробирке с кровью.
Тремя годами позже Giles и соавт. [18] идентифицировали антитела, откры-вавшие антиген Co b, антитетичный Co a.
использованием сыворотки анти-Co a обследованы 1706 американских не-гров, все оказались Со а + (Race, Sanger [45]). Среди 13 460 европейцев (жите-лей стран Северной Европы, США и Канады), обследованных разными автора-
ми, выявлены 27 человек Со(а −) (Heisto и соавт. [19], Lewis и соавт. [30], Race, Sanger [45]), Smith и соавт. [53], (Wray, Simpson [58]). Частота антигена Со а, та-
ким образом, составила 99,8 %.
Сыворотка анти-Co b использована при обследовании 799 испанцев, прожи-вающих в Майами (Issitt и соавт. [23]), 100 канадских индейцев племени кри (Lucciola и соавт. [31]) и 2244 японцев (Nagao и соавт. [38]. Частота выявления антигена Co b в указанных популяциях составила 4,6 %, 2 % и 0,22 % соответ-ственно.
Из 5186 доноров, обследованных Giles и соавт. [18] в Англии, Lewis и соавт.
193. в Канаде, Case [7], Brackenridge и соавт. [5] в Австралии и Новой Зеландии,
443 (8,5 %) имели фенотип Co(b + ) (табл. 17.2).
Результаты посемейных исследований [19, 30, 45] свидетельствуют, что гены Co a и Co b наследуются кодоминантно.
Описаны две семьи с необычным характером наследования генов Co a и Co b (Moulds и соавт. [35], Swanson, Eckman [55]). Количественное исследо-
вание антигенов эритроцитов членов этих семей путем титрования антител
741
анти-Co a и анти-Со3 показало, что они несли по одной дозе указанных анти-генов, что, по-видимому, было обусловлено передачей от одного из родителей молчащего аллеля Со.
Таблица 17.2
Дата: 2019-02-24, просмотров: 410.