Антигены Scianna расположены на гликопротеине, известном как HERMAP (human erythroid membarne associated protein – протеин, ассоциированный с мембраной эритроидных клеток человека). Синтез этого гликопротеина контро-лирует ген ERMAP, картированным на коротком плече хромосомы 1 в позиции 1р34.1. Ген ERMAP включает 11 экзонов протяженностью 19 пн (рис. 15.1).
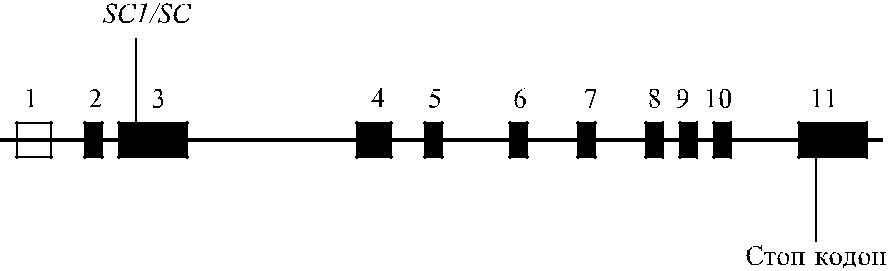
Рис. 15.1. Строение локуса ERMAP.
Протеин HERMAP состоит из 475 аминокислот (рис. 15.2) и представлен экстрацеллюлярным, трансмембранным и интрацеллюлярным доменами, вклю-чающими соответственно 157, 20 и 198 аминокислот (рис. 15.3). Его мол. мас-са 60–68 кДа.
| MEMASSAGSW LSGCLIPLVF LRLSVHVSGH AGDAGKFHVA LLGGTAELLC | 50 |
| PLSLWPGTVP KEVRWLRSPF PQRSQAVHIF RDGKDQDEDL MPEYKGRTVL | 100 |
| VRDAQEGSVT LQILDVRLED QGSYRCLIQV GNLSKEDTVI LQVAAPSVGS | 150 |
| LPSSAVALAV ILPVLVLLIM QCLCLIWKQR RAKEKLLYEH VTEVDNLLSD | 200 |
| HAKEKGKLHK AVKKLRSELK LKRAAANSGW RRARLHFVAV TLDPDTAHPK | 250 |
| LILSEDQRCV RLGDRRQPVP DNPQRFDFVV SILGSEYFTT GCHYWEVYVG | 300 |
| DKTKWILGVC SESVSRKGKV TASPANGHWL LRQSRGNEYE ALTSPQTSFR | 350 |
| LKEPPRCVGI FLDYEAGVIS FYNVTNKSHI FTFTHNFSGP LRPFFEPCLH | 400 |
| DGGKNTAPLV ICSELHKSEE SIVPRPEGKG HANGDVSLKV NSSLLPPKAP | 450 |
| ELKDIILSLP PDLGPALQEL KAPSF | 475 |
Рис. 15.2. Аминокислотная последовательность протеина HERMAP.
Молекулярная основа нулевого фенотипа Scianna разнородна (см. табл. 15.4).
В результате мутаций или делеции генетического материала у лиц с нулевым фенотипом (Sc: −1, −2, −3) синтезируемый протеин HERMAP утрачивает анти-генные эпитопы, распознаваемые антителами Scianna. Фенотипически это про-является в отсутствии всех без исключения антигенов указанной системы. Как уже отмечено выше, лица с фенотипом Scnull могут быть аллоиммунизированы часто встречающимся антигеном Sc3.
Полагают, что гликопротеин Scianna выполняет в эритроцитах функцию мо-лекул клеточной адгезии.
724
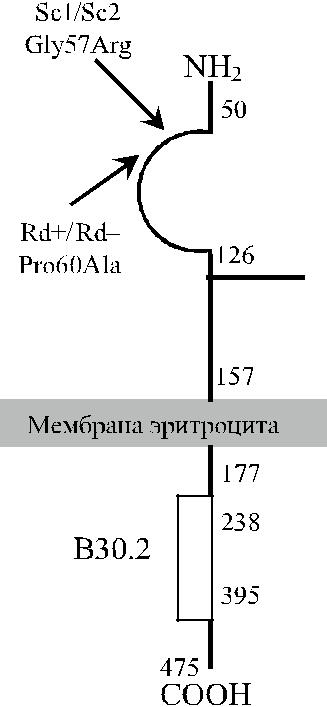
Рис. 15.3. Строение про-
теина HERMAP, несущего
антигены Scianna.
| Таблица 15.4 | |||||
Молекулярная основа полиморфизма антигенов Scianna
Scianna null
аминокислот
3
рамки считывания;
Список литературы
с Anderson C., Hunter J., Zipursky A. et al. An antibody defining a new blood group antigen, Bu a // Transfusion. – 1963. – V. 3. – P. 30–33.
с Banks J., Poole J., Ligthart P.C., Saez M. A complex serological investigation involving antibodies to two high frequency antigens [Abstract] // Transfus. Med. – 1998. – V. 8 (Suppl.). – P. 28.
с Calkovska Z. Mitteilung uber zwei weitere Familien mit einem Vorkommen von Bu a // Folia Haematol. – 1974. – V. 101. – S. 661–666.
с Daniels G. Effect of enzymes on and chemical modifications of high-frequency red cell antigens // Immunohematology. – 1992. – V. 8. – P. 53–57.
725
а Daniels G.L. Human Blood Groups. – 2-nd ed. – Oxford: Blackwell Science, 2002. – 560 p.
а DeMarco M., Uhl I., Fields L. et al. Hemolytic disease of the newborn due to the Scianna antibody, anti-Sc2 // Transfusion. – 1995. – V. 35. – P. 58–60.
а Devine P., Dawson F.E., Motschman T.L. et al. Serologic evidence that Scianna null (Sc:−1−2) red cells lack multiple high-frequency antigens // Transfusion. – 1988. – V. 28. – P. 346–349.
а Dunstan R.A. Status of major red cell blood group antigens on neutrophilis, lymphocytes and monocytes // Br. J. Haematol. – 1986. – V. 62. – P. 301–309.
а Flegel W.A., Che Q., Reid M. et al. SCER and SCAN: two novel high-prevalence antigens in the Scianna blood group system // Transfusion. – 2005. – V. 45. – P. 1940–1946.
а Funfhausen G., Gremplewski K. Die Verteilung des Blutgruppenantigens Bu a in Berlin //
Ärztl. Fortbild. Qualitatssich. – 1967. – V. 61. – S. 769.
а Gale S.A., Rowe G.P., Northfield F.E. Application of a microtitre plate antiglobulin technique to determine the incidence of donors lacking high frequency antigens // Vox Sang. – 1988. –
54. – P. 172–173.
В Hue-Roye K., Chaudhuri C A., Velliquette R.W. et al. STAR: a novel high-prevalence antigen in the Scianna blood group system // Transfusion. – 2005. – V. 45. – P. 245–251.
В Issitt P.D., Anstee D.J. Applied Blood Group Serology. – 4-th ed. – Durham, NC, USA: Montgomery Sc. Publ., 1998. – 1208 p.
В Kaye T., Williams E.M., Garner S.F. et al. Anti-Sc1 in pregnancy // Transfusion. – 1990. –
30. – P. 439–440.
В Lewis M., Chown B., Kaita H. On the blood group antigens Bu a and Sm // Transfusion. – 1967. – V. 7. – P. 92–94.
В Lewis M., Chown B., Kaita H., Griffits J. A possible relationship between the blood group antigens Sm and Bu a // Amer. J. Hum. Genet. – 1964. – V. 16. – P. 254–255.
В Lewis M., Chown B., Kaita H., Philipps S. Further observations on the blood group antigen Bu a // Am. J. Hum. Genet. – 1964. – V. 16. – P. 256–260.
В Lewis M., Kaita H. Genetic linkage between the Radin and Rh blood group loci // Vox Sang. – 1979. – V. 37. – P. 286–289.
В Lewis M., Kaita H., Chown B. Scianna blood group system // Vox Sang. – 1974. – V. 27. –
О 261–264.
У Lewis M., Kaita H., Philipps S. et al. The position of the some 1 loci // Ann. Hum. Genet. – 1980. – V. 44. – P. 179–184.
У Lundsgaard A., Jensen K.G. Two new examples of anti-Rd: a preliminary report on the frequency of the Rd(Radin) antigen in the Danish population // Vox Sang. – 1968. – V. 14. –
452–457.
У McCreary J., Vogler A.L., Sabo B. et al. Another minus – minus phenotype: Bu(a −)Sm–. Two examples in one family [Abstract] // Transfusion. – 1973. – V. 13. – P. 350.
У McDowell M.A., Stocker I., Nance S., Garratty G. Autoanti-Sc1 associated with autoimmune hemolytic anemia [Abstract] // Transfusion. – 1986. – V. 26. – P. 578.
У Mollison P.L., Engelfriet P., Contreras M. Blood Transfusion in Clinical Medicine. –10-th ed. – Oxford: BSP, 1997. – 1033 p.
У Nagao N., Tomita T., Okubo Y., Yamaguchi H. Low frequency antigen, Do a, Co b, Sc2 in Japanese (Abstract) // 24-th Cong. Int. Soc. Blood Transfus. – 1996. – P. 145.
У Nason G., Vengelen-Tyler V., Cohen N. et al. A high incidence antibody (anti-Sc3) in the serum of Sc:-1,-2 patient // Transfusion. – 1980. – V. 20. – P. 531–535.
У Noades J.E., Corney G., Cook P.J.L. et al. The Scianna blood group lies distal to uridine monophospate kinase on chromosome 1p // Ann. Hum. Genet. – 1979. – V. 43. – P. 121–132.
У Owen I., Chowdhury V., Reid M.E. et al. Autoimmune hemolytic anemia associated with anti-Sc1 // Transfusion. – 1992. – V. 32. – P. 173–176.
У Peloquin P., Moulds M., Keenan J., Kennedy M. Anti-Sc3 as an apparent autoantibody in two patient [Abstract] // Transfusion. – 1989. – V. 29. – 49S.
726
с Pierse S.R., Orr D.I., Brown P.J., Tilman G. A serum-reactive / plasma-nonreactive antibody with Scianna specificity [Abstract] // Transfusion. – 1989. – V. 29. – 36S.
с Race R.R., Sanger R. Blood Groups In Man., 6th ed., Oxford: Blackwell Scientific Publications, 1975.
с Rausen A.R., Rosenfield R.E., Alter A.A. et al. A ‘new’ infrequent red cell antigen, Rd(Radin) // Transfusion. – 1967. – V. 7. – P. 336–342.
с Schmidt R.P., Griffits J., Northman F.F. A new antibody, anti-Sm, reacting with a high incidence antigen // Transfusion. – 1962. – V. 2. – P. 338–340.
с Seyfried H., Frankowska K., Giles C.M. Further examples of anti-Bu a found in immunized donors // Vox Sang. – 1966. – V. 11. – P. 512–516.
с Spring F.A. Characterization of blood-group-active erythrocyte membrane glycoproteins with human antisera // Transfus. Med. – 1993. – V. 3. – P. 167–178.
с Spring F.A., Herron R., Rowe G. An erythrocyte glycoprotein of apparent 60000 expresses the Sc1 and Sc2 antigens // Vox Sang. – 1990. – V. 58. – P. 122–125.
с Tregellas W.M., Holub M.P., Moulds J.J., Lacey P.A. An example of autoanti-Sc1 demonstrable in serum but not in plasma [Abstract] // Transfusion. – 1979. – V. 19. – P. 650.
с Woodfield D.G., Giles C., Poole J. et al. A further null phenotype (Sc:–1,–2) in Papua New Guinea [Abstract] // 19-th Cong. Int. Soc. Blood Transfus. – 1986. – P. 651.
с Yung C.H., Chow M.P., Hu H.Y. et al. Blood group phenotypes in Taiwan // Transfusion. – 1989. – V. 29. – P. 233–235.
727
Глава 16.
Система Dombrock
До 1995 г. к системе Dombrock (Домброк) относили два антитетичных анти-гена – Do a и Do b. В 1995 г. Banks и соавт. [1] установили, что у лиц, лишенных антигенов Do a и Do b, отсутствуют также еще три антигена – Gy a, Hy и Jo a – и они имеют фенотип Do(a −b −)Gy(a −)Hy −Jo(a −). Столь очевидная фенотипи-ческая связь позволила расширить систему Dombrock до пяти единиц. В нее были включены часто встречающиеся антигены Gy a (Gregory), Hy (Holley) и Jo a(Joseph), получившие обозначения DO3, DO4 и DO5 в номенклатуре ISBT. Антигены Do a и Do b получили обозначения DO1 и DO2 (табл. 16.1).
| Таблица 16.1 | ||||||
Антигены Dombrock
Обозначение
Примечание
традиционное
ISBT
европеоидов, %
Hy (Holley)
Jo a (Joseph)
DOYA
Антигены Do a, Do b и Jo a полностью развиты к моменту рождения (Swanson
и соавт. [45], Molthan и соавт. [23], Laird-Fryer и соавт. [18], Jensen и соавт. [14]).
Антигены Gy a и Hy, наоборот, на эритроцитах новорожденных выражены слабо
(Clark и соавт. [6], Moulds и соавт. [26]).
Обработка эритроцитов папаином или фицином повышает серологическую активность антигенов Dombrock, поэтому антиглобулиновый тест с клетками, предварительно обработанными указанными ферментами, является оптималь-ным, особенно при определении антигенов Do a и Do b.
Вещество Dombrock разрушается трипсином, химотрипсином, проназой и сульфгидрильными редуцентами. Сиалидаза подобного эффекта не оказывает
(Banks и соавт. [1], Brown [4], Spring и соавт. [42, 43]).
728
Антигены Dombrock расположены на гликопротеине, связанном с гликозил-фосфадитилинозитолом (ГФИ). Этот пептид относится к группе аденозинди-фосфатрибозилтрансфераз.
Генный локус DO картирован на хромосоме 12 в позиции 12р13.2-12.1.
Do a и Do b
1965 г. Swanson и соавт. [45] обнаружили у европейки по фамилии Dombrock антитела, реагирующие с эритроцитами примерно 64 % произвольно выбранных доноров. Авторы показали, что антиген, обозначенный ими Do a, не связан с другими антигенами эритроцитов и может быть отнесен к новой, ранее неизвестной системе.
1973 г. Molthan и соавт. [23] нашли антитела, открывающие антитетичный антиген – Do b.
Данные о частоте антигенов Dombrock у различных народов неполны, по-скольку получены, в основном, с использованием сывороток анти-Do a. Чаще всего антиген Do a встречается у европеоидов и негроидов, реже – у монголои-дов (табл. 16.2, 16.3). Полученная на основе этих данных расчетная частота ге-нотипов Do a / Do a, Do a / Do b и Do b / Do b составила соответственно 0,1764; 0,4872
20. 0,3364.
| Таблица 16.2 | |||||||
| Частота антигена Do a и генов Do a и Do b у разных народов | |||||||
| Количество | Частота антиге- | Частота генов | |||||
| Популяция | на Do | a | Источник | ||||
| обследованных | |||||||
| абс. ч. | % | Do a | Do b | ||||
| Жители стран | 755 | 501 | 66,36 | 0,4200 | 0,5800 | [45, 48, | |
| Северной Европы | 49] | ||||||
| Белые американцы | 391 | 250 | 63,94 | 0,3395 | 0,6005 | [30] | |
| Жители стран | 700 | 446 | 63,71 | 0,3976 | 0,6024 | [19] | |
| Северной Америки | |||||||
| Американские негры | 161 | 89 | 55,28 | 0,3313 | 0,6687 | [30] | |
| Американские негры | 76 | 34 | 44,74 | 0,2566 | 0,7434 | [48] | |
| Японцы | 760 | 179 | 23,55 | 0,1257 | 0,8743 | [27, 28] | |
| Жители Таиланда | 423 | 57 | 13,48 | 0,0698 | 0,9302 | [5] | |
Исследование эритроцитов более 2500 членов канадских, израильских, япон-ских, негритянских и других семей с помощью сывороток анти-Do a показало, что во всех случаях независимо от расовой принадлежности ген Do a проявлял себя как аутосомно-доминантный признак (Tippett и соавт. [48, 49], Lewis и со-авт. [19], Polesky, Swanson [30]). У родителей Do(a + ) × Do(a −) частота ожидае-мых и частота фактических фенотипов детей по антигену Do a совпадали.
729
Таблица 16.3
Частота фенотипов Dombrock
| Сочетание антигенов | Частота (%) среди | |||||
| Do a | Do b | Gy a | Hy | Jo a | европеоидов | негроидов |
| + | – | + | + | + | 18 | 11 |
| + | + | + | + | + | 49 | 44 |
| – | + | + | + | + | 33 | 45 |
| – | – | – | – | – | Редко | 0 |
| – | сл | сл | – | – | 0 | Редко |
| сл | сл | + | сл | – | 0 | Редко |
Примечание: « + » – антиген присутствует; « – » – антиген отсутствует; сл – антиген слабо выражен.
Gy a
1967 г. (Swanson и соавт. [46]) описали американскую семью чешского происхождения, в которой у 4 из 7 детей отсутствовал часто встречающийся ан-тиген Gy a (Gregory). В сыворотке крови пропозиты и ее сестры присутствовали антитела анти-Gy a. Первая имела пять беременностей, вторая – две.
другой семье, также чешского происхождения, было две женщины Gy(a −), содержавшие анти-Gy a-антитела. Обе женщины имели повторные беременно-
сти (Race, Sanger [32]).
Еще 6 индивидов Gy(a −) выявлены в английской семье, имевшей, как пола-гают Clark и соавт. [6], романское происхождение. У 4 сестер обнаружены анти-Gy a-антитела, у 2 их братьев они отсутствовали. Сестры имели многократные беременности.
Okubo и соавт. [29] описали 6 японских женщин Gy(a −). Редкий фенотип вы-явлен в связи с обнаружением у них анти-Gy a-антител.
Mak и соавт. [20] наблюдали мужчину Gy(a −) (китайца из Гонконга), имев-шего анти-Gy a-антитела, вероятно, аллоиммунной природы: 40 лет назад ему перелили кровь.
Антиген Gy a присутствует практически у всех людей. Лица Gy(a −) не были обнаружены при обследовании 9350 японцев (Okubo и соавт. [29]) и 10 145 аме-риканцев (Swanson и соавт. [46]). Среди негроидов Фенотип Gy(a −) не выявлен.
Из большого числа обследованных (4530 белых американцев, 735 чехов, 683 белых южноафриканца, 846 американских негров, 1023 черных южноафриканца, 633 южноафриканских индуса, 1679 американских индейцев) только один чело-век – европеец по происхождению – имел фенотип Gy(a −) и два индейца племени апачи имели фенотип Gy(a + w). Остальные обследованные были Gy(a + ).
Ген, формирующий фенотип Gy(a −), наследуется по рецессивному типу. Описаны супружеские пары Gy(a + ) × Gy(a + ), имевшие детей Gy(a −) (Swanson
В соавт. [46], Clark и соавт. [6], Okubo и соавт. [29], Massaquoi [21]). Родители,
730
как правило, были кровными родственниками (Swanson и соавт. [46], Okubo и соавт. [29], Massaquoi [21]).
На эритроцитах Gy(a −) отсутствуют антигены Ну и Jo a (Moulds и соавт. [26], Laird-Fryer и соавт. [18]), а также Do a и Do b. Подобная фенотипическая зависи-мость указывала на то, что перечисленные антигены могут входить в одну систему.
Hy
Первый образец сыворотки, содержащей анти-Ну-антитела, получили Schmidt и соавт. [40]. Позднее были найдены другие образцы антител указан-ной специфичности (Moulds и соавт. [26], Beattie, Castillo [3], Hsu и соавт. [12]). Носителями антител во всех случаях были негроиды, имевшие фенотип Ну −.
Moulds и соавт. [26] отметили фенотипическую связь антигенов Ну и Gy a. Негроиды Ну − имели слабый антиген Gy a [фенотип Ну −Gy(a + w)], тогда как у европеоидов Ну − и монголоидов Ну − антиген Gy a отсутствовал.
Индивиды Ну −Gy(a −) вырабатывали анти-Gy a-антитела, индивиды Ну −Gy(a + w) – анти-Ну-антитела.
Сыворотки анти-Gy a не содержали антител анти-Ну. Троекратная адсорбция сывороток анти-Gy a эритроцитами Ну −Gy(a + w) полностью истощала активность анти-Gy a-антител. Элюаты с указанных эритроцитов вели себя в серологических реакциях так же, как антитела исходных неадсорбированных сывороток анти-Gy a.
Все лица Ну −Gy(a + w) были Do(a −b + w), антиген Do a у них отсутствовал, ан-тиген Do b был выражен слабо (Banks и соавт. [1]).
с 7 лиц Ну −Gy(a + w), обследованных с использованием молекулярно-биологических методов, была найдена замена нуклеотида G 232 T в экзоне 2, ведущая к аминокислотной замене Gly 108 Val (Rios и соавт. [37, 38]). У указан-ных индивидов обнаружена мутация C 898 G в экзоне 3, которая приводила к аминокислотной замене Leu 300 Val и, соответственно, уменьшала экспрессию антигенов Gy a и Do b.
Jo a
выявлении антигена Jo a (Joseph) впервые сообщили Jensen и соавт. [14], которые нашли антитела, идентифицировавшие этот антиген у 2 пациентов – американских негров. Обе сыворотки реагировали со всеми исследованными эритроцитами.
Третий образец анти-Jo a-антител обнаружили Morel и соавт. [24] у больного
и серповидно-клеточной анемией.
Все три носителя анти-Jo a-антител получали многократные гемотрансфузии. Laird-Fryer и соавт. [18] выявили у 5 негритянок антитела, обозначенные как анти-Jc a. Позднее было показано, что эти антитела и антитела анти-Jo a реагиру-
ют с одним и тем же антигеном (Weaver и соавт. [52]).
Частота встречаемости антигена Jo a очень высока. Jensen и соавт. [14] при об-следовании 3000 жителей Нью-Йорка, преимущественно европеоидов, а также
731
7689 американских негров не нашли ни одного человека с фенотипом Jo(a −). Позднее было установлено, что антиген Jo a отсутствует на эритроцитах лиц
Hy −Gy(a −) и Hy −Gy(a + w), что свидетельствовало о фенотипической связи этих антигенов и возможной принадлежности к одной групповой системе. Вместе с тем все лица, имевшие анти-Jo a-антитела, были Gy(a + )Hy + Jo(a −) (Laird-Fryer и соавт. [18], Weaver и соавт. [51], Brown и соавт. [4]). Тем самым было показано, что антигены Gy a, Ну и Jo a полностью различаются между собой.
Антиген Ну несколько отличался от антигена Jo a. Если антиген Jo a отсут-ствовал на эритроцитах Hy −Gy(a −) и обнаруживался только на эритроцитах Hy + Gy(a + ), то антиген Ну присутствовал на эритроцитах и Gy(a + )Jo(a + ), и Gy(a + ) Jo(a −) (Spring и соавт. [43]).
Weaver и соавт. [51] нашли 4 человек Ну −Gy(a −)Jo(a −).
Banks и соавт. [1] выявили пять человек Jo(a −)Do(a + wb + w) негров, одного испанца Jo(a −)Do(a + wb −).
Дата: 2019-02-24, просмотров: 375.