Сыворотки с антитела-
Реакция эритроцитов, имеющих фенотип
ми к антигенам
обычновстречающимся
c
–
–
( + )
–
–
–
–
редковстречающимся
частовстречающимся
Примечание. Реакция: + положительная, − отрицательная, ( + ) слабоположительная.
−D −
Первый из выявленных фенотипов делеций ( −D −) обнаружили Race, Sanger
и Selwyn в 1951 г. Затем были описаны другие случаи гомо- и гетерозигот −D − [149, 229, 480, 502, 556, 592, 725]. Последний случай опубликован в 1991 г. Moores и соавт. (Humen Hered., 1991, V. 41, P. 295).
Эритроциты −D − не содержат антигенов С, с, Е и е. При этом антигены D, G, Rh total и Rh39 сильно выражены; часто встречающиеся антигены: Hro, Nou, Dav и др. – отсутствуют (см. табл. 4.22).
244
В России фенотип −D − обнаружен в 1985 г. В.А. Мороковым [82] и подроб-но исследован Т.М. Пискуновой с соавт. [87]. Описанный случай представляет собой классический пример фенотипа −D −.
Наблюдение. Беременная З., 35 лет, В(III) Rh +, диагноз – неразвивающаяся бе-ременность 24 недели. В анамнезе 1 гемотрансфузия и 6 беременностей, из которых первые 2 закончились рождением здоровых детей, 3-я и 4-я – искусственным преры-ванием, 5-я – внутриутробной гибелью плода, 6-я беременность, неразвивающаяся,
послужила показанием к экстирпации матки. Эритроциты З. содержали антиген D, но не содержали антигенов C, c, E, e и C W.
Бабушка, отец и мать З. имели гаплотип −D − в гетерозиготном варианте. Всего в семье обнаружено 3 гомозиготы и 6 гетерозигот −D − (рис. 4.10).
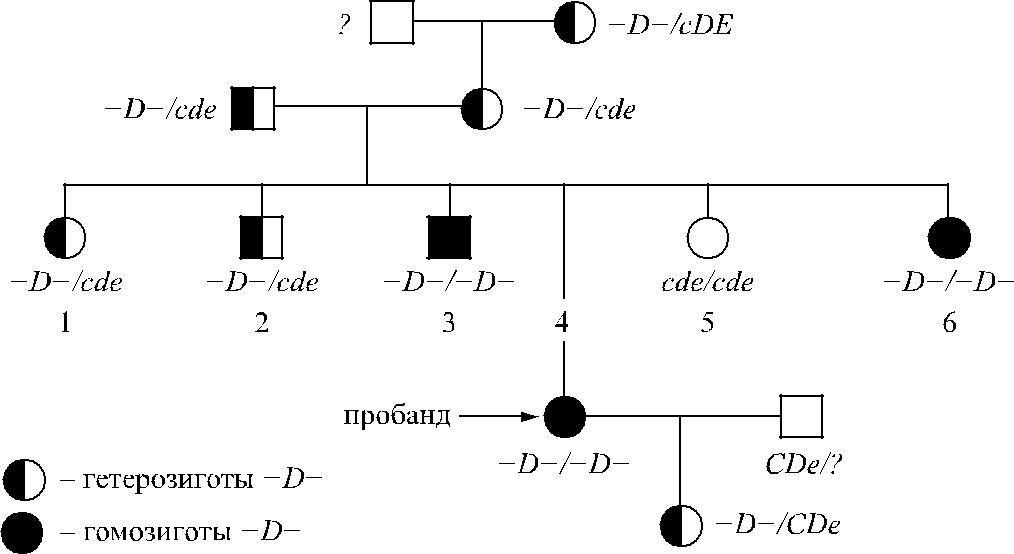
Рис. 4.10. Распределение гаплотипа −D− в семье З.
Эритроциты гетеро- и гомозигот −D − проявляли необычайно высокие агглютина-бельные свойства. Титр сывороток анти-D при титровании этими эритроцитами соот-ветствовал 1 : 256–1 : 1024, в то время как с эритроцитами доноров контрольной группы титр антител не превышал 1 : 64 (табл. 4.23). Наиболее выраженная агглютинабельность эритроцитов отмечена у гомозигот −D −/ −D −. Результаты этих экспериментов совпада-ют с данными других авторов, свидетельствующими о том, что эритроциты −D − содер-жат повышенное количество D-несущих полипептидов (см. Экспрессия антигена D).
Сыворотка З. реагировала со всеми образцами стандартных эритроцитов не-зависимо от сочетания Rh-антигенов, поэтому подобрать ей совместимую кровь среди доноров оказалось невозможным. С собственными эритроцитами и эри-троцитами одного из братьев и одной из сестер сыворотка З. не реагировала.
В методе солевой агглютинации положительный результат наблюдали только
с эритроцитами, содержащими антиген С, что указывало на присутствие в сы-воротке полных анти-С-антител.
245
Таблица 4.23
Реагирование −D − эритроцитов членов семьи З. с сывороткой анти-D
| Титр анти-D-антител с | |||
| Член семьи | Генотип | эритроцитами членов семьи | |
| и доноров контрольной группы | |||
| Бабушка | −D −/cDE | 1 : 512 | |
| Мать | −D −/cde | 1 : 512 | |
| Сестра | −D −/cde | 1 : 256 | |
| Брат | −D −/cde | 1 : 256 | |
| Брат | −D −/ −D − | 1 : 1024 | |
| Пробанд | −D −/ −D − | 1 : 1024 | |
| Сестра | cde / cde | 0 | |
| Сестра | −D −/ −D − | 1 : 1024 | |
| Доноры | CDe / cde | 1 : 32 | |
| CDe / CDe | 1 : 32 | ||
| (контрольная группа) | |||
| cDE / cDE | 1 : 64 | ||
Учитывая отсутствие в эритроцитах больной четырех других, типичных для системы резус минорных антигенов, авторы предположили, что сыво-ротка З. содержит полиспецифические антирезус-антитела, выработка кото-рых могла быть индуцирована многократными беременностями. Исследование крови мужа показало, что его фенотип CcDee мог соответствовать генотипу CDe / cde, CDe / CDe или cDe / Cde, следовательно, генотипы детей З. могли быть −D −/ cde, −D −/CDe или −D −/Cde, что с большой степенью вероятности могло привести к иммунизации женщины факторами С, с и е. Имеющаяся в анамнезе З. гемотрансфузия давала основание подозревать аллосенсибилизацию к анти-гену Е. Пробы с дифференциальной адсорбцией позволили идентифицировать антитела, содержащиеся в сыворотке З., как анти-С полные с титром 1 : 256, не-полные анти-с с титром 1 : 2048 и неполные анти-е с титром 1 : 128.
Резюмируя описанное В.А. Мороковым наблюдение, уместно подчер-кнуть, что антитела к минорным антигенам С, с и е встречаются относитель-но редко и имеют низкий титр. На этом фоне способность продуцировать антитела одновременно к обоим антитетичным антигенам – С и с – и к анти-гену е (hr") в столь высоком титре подчеркивает особый иммунологический статус гомозигот −D −/ −D −, характеризующийся повышенной чувствитель-ностью к недостающим факторам Rh, которые являются для них сильным иммуногеном.
*D*
Эритроциты *D* обнаружены Contreras и соавт. [237]. Они содержат по-вышенное количество антигена D и в серологических реакциях проявля-ют себя подобно эритроцитам −D −, однако в отличие от последних содер-жат антигены Evans (Rh37) и Dav (Rh47). Особенности фенотипа *D*, от-личающие его от других фенотипов делеций, представлены в табл. 4.22.
246
Фенотип *D*, как и другие фенотипы Rh-делеций, не содержит общих, ши-роко распространенных Rh-антигенов, присущих нормальному Rh-комп лексу (Hro, Hr B , Sec и MAR), а также большинства редких Rh-антигенов (Go a, Tar, FPTT и др.).
Миссис Helen Dav, у которой были найдены эритроциты *D*, имела антите-
ла, реагирующие с некоторыми образцами эритроцитов от лиц с Rh-делециями,
а ее эритроциты реагировали с некоторыми сыворотками от указанных лиц. Анализ особенностей реагирования эритроцитов миссис Dav привел к откры-тию одноименного антигена (см. Dav).
DC W −
Эритроциты DC W −, описанные в 1957 г. Gunson, Donohue [327], характери-зуются наличием антигена G и отсутствием большинства часто встречающихся антигенов Rh-комплекса (см. табл. 4.22). Антигены С, с, Е и е также отсутству-ют, на что указывает знак минус в обозначении фенотипа. Как было показано Tippett [657], антиген C W в эритроцитах DC W − выражен слабее, чем в эритроци-тах с нормальным фенотипом DCC We.
Dc −
Первое сообщение о фенотипе, обозначенном Dc −, относится к 1960 г. (Tate
и соавт. [649]), затем появились другие публикации [258, 440, 724]. Как было установлено, эритроциты Dc − не содержат антигенов С, Е и е, но отличают-ся большой вариабельностью в отношении количества антигена с (hr'). Так, по данным Yamaguchi и соавт. [724], экспрессия антигена с (hr') на эритроцитах Dc − снижена, антиген f на них отсутствует.
Tate и соавт. [649] нашли, что, несмотря на ослабленный антиген с (hr') и отсутствие антигена e (hr"), эритроциты Dc − содержат антиген f, но в осла-бленной форме. Отдельные образцы эритроцитов Dc −, содержали некоторое количество антигена e (hr"). Такой фенотип более соответствовал написанию Dc(e), т. е. Dc со слабым e (hr"), или Dc((e)), т. е. Dc с исчезающим количе-
ством e (hr").
Leyshon и соавт. [440] привели данные о нормальной выраженности анти-гена с (hr') и f (се) на эритроцитах Dc −. Однако Spielman и соавт. [Vox Sang., 1974, V. 27, P. 473] описали случай ГБН, вызванный анти-f-антителами, у жен-щины, которая имела генотип CDe / Dc-, что вступает в противоречие с данными
Leyshon и соавт.
Многие авторы сходятся во мнении, что количество антигена D в эритроци-тах Dc − выше, чем в обычных эритроцитах D +.
Tessel и соавт. [650] описали фенотип Dc − негра и негритянки, не свя-занных родством. Эритроциты мистера J.W. содержали повышенное ко-личество антигена D, уменьшенную по сравнению с нормой дозу анти-генов с (hr') и f, а также слабый антиген e (hr"). Эритроциты миссис F.J.
247
содержали повышенное количество антигена D, уменьшенную дозу антиге-на с (hr'), но не содержали антигенов f и e (hr"). Эритроциты мистера J.W. реагировали с антителами анти-Hro / Hr, имевшимися в сыворотке крови миссис F.J. и двух гомозигот −D −/ −D −, а также с двумя из трех сывороток анти-nl. Эритроциты миссис F.J. не реагировали с сыворотками анти-Hro / Hr и анти-nl. Авторы пришли к выводу, что ген Dc − гетерогенен и может ко-дировать 2 фенотипа Dc −, один из которых содержит антиген f, трудновы-являемый антиген е, антигены Hro и Hr, а другой фенотип Dc − не содержит перечисленных антигенов.
D IV (C) −
В 1969 г. группой исследователей [593], включая классиков современной им-муносерологии Sanger, Tippett и Salmon, был описан единственный известный до настоящего времени человек с генотипом D IV (C) −/D IV (C) −. Людей, гетеро-зиготных по гаплотипу D IV (C) − обнаружено несколько. Гаплотип D IV (C) − про-изводит парциальный антиген D категории IVa, антиген G и значительно умень-шенное количество антигена С.
В гомо- и гетерозиготном варианте генный комплекс D IV (C) − обеспечивает экспрессию редко встречающихся антигенов: Go a (Rh30) [593], Ro Har (Rh33), Riv (Rh45) и FPTT (Rh50) [257], а также часто встречающихся Rh-антигенов: Rh to-tal (Rh29), Rh ауто-С-подобный (Rh39), Dav (Rh47) и Nou (Rh44), которые не ко-
дируются другими гаплотипами Rh-делеций (см. табл. 4.22).
Антиген Nou был впервые обнаружен на эритроцитах гомо- и гетерозигот
D IV (C) − (см. Nou).
Хотя эритроциты D IV(C) − лишены антигена е (hr"), 3 из 10 сывороток анти-е давали реакции с этими эритроцитами, что указывает на продукцию некоторого количества антигена е (hr") или Се (rhi) гаплотипом D IV (C) −.
Rh null
Среди упомянутых выше фенотипов делеций ( −D −, Dc −, D IV(C) − и DC W −), при которых наблюдается выпадение 2 антигенов и более, фенотип Rh null зани-мает особое место. Он отличается отсутствием всех антигенов системы Rh-Hr, а также антигенов системы LW и Duffy, кодируемых генами, близко расположен-ными к локусу RH.
О первой находке необычного генотипа − − −/ − − − у аборигенки Австралии сообщили в 1961 г. Vos и соавт. [689]. В 1964 г. такой же генотип описали Levine
и соавт. [428] у белой американки. Двумя годами позже, в 1966 г., Ishimori, Hasekura [371] нашли нулевой Rh-фенотип у японского мальчика.
248
| Таблица 4.24 | ||||||||||
| Сводные данные о лицах Rh null | ||||||||||
| Пробанд | Расовая | Экспрессия | Сибсы | Родители – | ||||||
| ность | Rhnull | родственников | Rhnull | Rh | родственники | |||||
| принадлеж- | Тип | Rh-антигенов у | не | кровные | Источник | |||||
| null | ||||||||||
| 1 | Аборигены | P | Снижена | 0 | 0 | нд | Vos и соавт. [689] | |||
| Австралии | ||||||||||
| 2 | Белые, | Р* | Снижена | 0 | 3 | нет | Levine и соавт. | |||
| США | [427, 428] | |||||||||
| Ishimori, Hasekura. | ||||||||||
| 3 | Японцы | А | нд | 0 | 1 | да | [371], см. также | |||
| [338] | ||||||||||
| 4 | Белые, | Р* | В норме | 0 | 0 | да | Haber и соавт. [331], | |||
| США | см. также [153] | |||||||||
| 5 | Французы | Р | Снижена | 2 | 2 | да | Guevin и соавт. цит | |||
| по [544] | ||||||||||
| 6 | Белые, | *? | нд | 1 | 0 | нд | Sturgeon [638] | |||
| США | ||||||||||
| 7 | Белые, | Р | Снижена | 0 | 0 | да | Senhauser и соавт. | |||
| США | [602] | |||||||||
| 8 | Немцы | А?* | В норме | 1 | 0 | да | Seidl и соавт. [601] | |||
| Белые, | Polesky и соавт. | |||||||||
| 9 | Р | В норме | 2* | 0 | нет | [532], см. также | ||||
| США | ||||||||||
| [492] | ||||||||||
| 10 | Немцы | ?* | Снижена | 0 | 0 | да | Nagel и соавт. [501] | |||
| 11 | Чехи | Р | В норме | 0 | 1 | нд | Hrubisko и соавт. | |||
| [353] | ||||||||||
| 12 | Белые, | Р | В норме | 1 | 5 | да | Stevenson и соавт. | |||
| США | [629] | |||||||||
| 13 | Норвежские | А | нд | 1 | 8 | да | Oestgaard цит. по | |||
| лапландцы | [544] | |||||||||
| 14 | Шведы | Р | Снижена | 0 | 3 | нет | Müller цит. по [544] | |||
Примечание. Р – регуляторный тип Rhnull , А – аморфный, нд – нет данных, * лица Rhnull , в крови которых обнаружены антиэритроцитарные антитела.
По данным Race и Sanger [544], к 1975 г. было описано более 20 лиц Rhnull из
14 семей, отличавшихся расовой принадлежностью (табл. 4.24). К этой сводке можно добавить 11 случаев Rhnull, приведенных Daniels [244].
Issitt и Anstee [374] насчитали 33 случая Rhnull, 1 из которых обнаружен ими. Фенотип Rhnull у лиц, внесенных в табл. 4.24, диагностирован при различных обстоятельствах. В 1-м случае он был обнаружен при проведении популяцион-ных исследований у австралийских аборигенов. В других случаях (2, 4, 6, 8, 10, 11 и 12-м) – при выяснении специфичности антител, в том числе у больных тяже-лой формой гемолитической анемии (8, 11 и 12-м), а также при обследовании до-
норов, рожениц, новорожденных (3, 5, 7, 9, 13 и 14-м).
249
Родственники носителей Rhnull в некоторых случаях также имели нулевой фе-нотип или сниженную экспрессию Rh-антигенов.
По сводке Daniels [244], из 11 лиц, имевших дефицит Rh-антигенов, 8 были детьми от близкородственных браков. Из общего числа обследованных 6 чело-век японцы.
Относительно большое число лиц Rhnull среди японцев обусловлено, скорее всего, не тем, что в этой популяции высокая частота указанного фенотипа, а проведением широких популяционно-скрининговых исследований, в процессе которых возможно выявить людей со столь редким фенотипом.
Обозначение – – – / – – –, использованное Vos и Levine с соавторами [428, 689], было признано неточным, поскольку означает отсутствие генетического материала у носителей этого фенотипа. В действительности гены RH у таких людей часто присутствуют, но не проявляют себя фенотипически. По предло-жению Ceppellini фенотип с отсутствием Rh-антигенов получил название Rhnull. Обозначение – – – / – – – сохранилось для описания гена r=, который, как позд-нее выяснилось, представляет собой молчащий (не производящий продукта) ал-лель локуса RH.
Регуляторный и аморфный тип Rh null
Различают 2 типа Rhnull: регуляторный (супрессорный) и аморфный (отсут-ствие генетического материала).
Регуляторный тип формируется в результате взаимодействия генов RH, производящих соответствующие полипептиды, и генов X 1 r, X O r, которые управ-ляют функцией локуса RH. Ген X 1 r (доминантный) запускает нормальную про-дукцию Rh-полипептидов, а его редкий аллель X O r (рецессивный) блокирует их синтез.
Ген X 1 r широко распространен, поэтому большинство людей являются гомо-зиготами – X 1 r / X 1 r. При таком генотипе унаследованные от родителей гены RН функционируют нормально. При гетерозиготном варианте ( X 1 r / X O r) экспрес-сия антигенов, по данным одних авторов, не нарушена [331, 532], по данным других – снижена [353].
У людей, гомозиготных по супрессорному гену, X O r / X O r, присутствующие RН-гены не функционируют. На рис. 4.11 представлен пример наследования ре-
гуляторного типа Rhnull.
Аморфный тип Rhnull обусловлен отсутствием генетического материала, ко-дирующего продукцию Rh-полипептидов, по-видимому, вследствие делеции.
Получены данные, которые можно трактовать в пользу существования мол-чащего гена r= [371]. Пример наследования аморфного типа Rhnull представлен на рисунке 4.12. Родители пробандов – сибсы. Их фенотипы cDe и CDe создают видимость гомозиготности по генам с и С.
Судя по фенотипу детей, оба родителя были гетерозиготами: один, вероятно, cDe / r=, второй – CDe / r=. Двое детей имели одинаковый генотип r=/r=. Трое
250
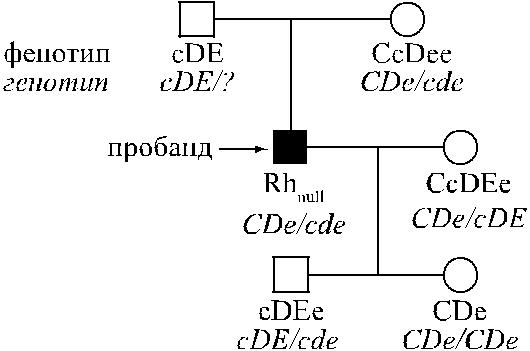
Рис. 4.11. Наследование регуляторного типа Rhnull по Hrubisko и соавт. [353]. Родители пробанда, по-видимому, гетерозиготы – X 1 r/X O r. Пробанд Rhnull передал детям нормальные гаплотипы cde и CDe.
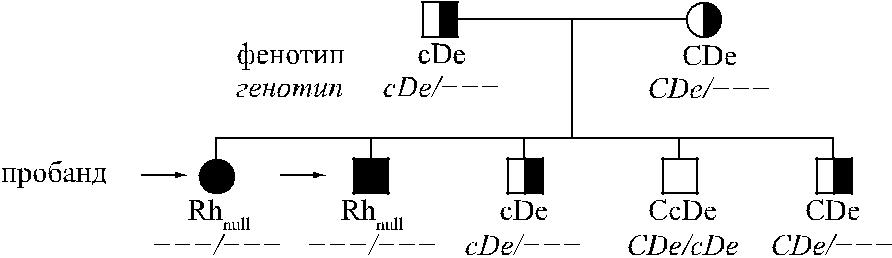
Рис. 4.12. Наследование аморфного типа Rhnull (Oestgaard, цит. по [544]).
Родители пробандов, по-видимому, гетерозиготы по молчащему гену r=.
других детей имели генотипы cDe / r=, CDe / r= и CDe / cDe. В приведенном при-мере недостает представителей третьего поколения, фенотипы которых могли бы подтвердить, что от родителей с аморфным типом Rhnul возможно рождение детей с таким же аморфным типом.
Обследование малого числа поколений в семьях не позволяет идентифи-цировать аморфный тип Rhnull, поскольку трудно получить убедительные до-казательства того, что конкретный обследуемый человек генетически r=/r=, а не X O r / X O r. Генотип X 1 r / X O r также может приводить к уменьшению экспрес-сии антигенов Rh. Однако совокупность фактов, полученных при обследовании большого количества семей, позволила исследователям сделать вывод, что на-следование Rhnull через молчащий ген r= вполне возможно.
Из 14 обладателей фенотипа Rhnull (см. табл. 4.24) у 9 был обнаружен регуля-торный тип Rhnull, у 3 – аморфный, у 2 – тип Rhnull не установлен.
Среди упомянутых выше 33 обследованных лиц Rhnull 20 имели регулятор-ный тип, 3 – аморфный тип, у остальных 10 человек генетическую основу нуле-вого фенотипа установить не удалось. Последующие многочисленные исследо-вания подтвердили, что оба типа, регуляторный и аморфный, встречаются край-не редко, причем один из них, регуляторный, преобладает [151, 153, 331, 353, 532]. В некоторых случаях семейные исследования не могли выявить генетиче-скую основу фенотипа Rhnull [501, 647].
251
Определенную роль в происхождении Rhnull, как и других фенотипов деле-ций, играют браки между родственниками, способствующие накоплению гомо-зигот по депрессивным и молчащим аллелям. Как видно из приведенных выше данных, родители лиц Rhnull более чем в половине случаев являлись кровными родственниками.
Молекулярно-генетические исследования, проведенные Сherif-Zahar и со-авт. [210] и Huang и соавт. [357], выявили делецию гена RHD и 2 мутации, обу-словливающие Rhnull аморфного типа. Выявлена делеция 2 нуклеотидов в кодо-нах 322 и 323 экзона 7 гена RHCE с нуклеотидной заменой TCA → C. Такой ген производит укороченный протеин – 10 трансмембранных доменов из 398 ами-нокислот вместо 12 доменов из 417 аминокислот. Авторы полагают, что указан-ные изменения мешают взаимодействию Rh-протеина с Rh-ассоциированным гликопротеином (RhAG), в результате чего полноценный Rh-комплекс на по-верхности мембраны эритроцита не образуется.
и одном случае Rhnull Huang и соавт. [358] нашли 2 перемещения G → A в экзоне 6 гена RHAG (ассоциированного гликопротеина), которые приводили к замене Val 270 → Ile и Gly 280 → Arg. Другой случай Rhnull характеризовался трансверсией G → T в экзоне 9, которая вызывала замещение Gly 380 → Val в трансмембранном сегменте 12.
Антитела, образующиеся у лиц Rh null
Из 22 носителей Rhnull (пробандов и их родственников) 7 (30,2 %) содержа-ли антиэритроцитарные антитела (см. табл. 4.24), что свидетельствует о весьма высокой частоте аллоиммунизации лиц, лишенных антигенов резус. Антитела имели аллоиммунный характер (беременности, трансфузии), за исключением одного случая спонтанных антител у мужчины – донора крови.
1 пробанда имелись антитела анти-С и анти-е, у 2 антитела были слабыми
32. не идентифицировались.
Важная деталь: 4 из 7 сывороток (пробанды 4, 8, 10 и 1 из 2 сестер 9-го) со-держали антитела, реагирующие с эритроцитами всех фенотипов Rh, в том чис-ле −D −/ −D − и других Rh-делеций, за исключением эритроцитов Rhnull. Эти ан-титела, подробно изученные Haber и соавт. [331] и Bar-Shany и соавт. [153], по-лучили название anti-total-Rh, а выявляемый ими широко распространенный ан-тиген был обозначен как Rh total (Rh29).
38 родильниц Rhnull отмечены случаи умеренной и тяжелой ГБН, вызванной анти-Rh29-антителами. Тяжелый случай успешно купирован обменными транс-фузиями крови cde в течение первых 24 ч после рождения ребенка (Lubenko и соавт. [451]). В одном случае ГБН была связана с анти-Hro-антителами (Perez-Perez и соавт. [Am. J. Hematol., 1992, V. 40, P. 306]).
По-видимому, иммунный ответ у лиц Rhnull ограничивается образованием ан-тител к общим антигенам – Rh29 и Hro. Антитела к частным антигенам (С, е и др.) вырабатываются реже.
252
При искусственной иммунизации добровольцев эритроцитами Rhnull анти-тел к веществу Rhnull получить не удалось. Известна лишь одно сообщение, свидетельствующее о возможности существования таких антител . McGinnis и соавт. [471, 472] нашли у 4 больных анемией антитела IgM и IgG, которые они назвали антипрекурсорными. Сыворотки реагировали только с эритроцитами Rhnull. Со стандартными эритроцитами (не Rhnull) агглютинации не наблюдали, однако после обработки стандартных эритроцитов трипсином реакция все же происходила.
Повреждение других антигенов на эритроцитах Rh null
12. табл. 4.25 суммированы имеющиеся в литературе данные относительно изменения статуса некоторых групповых антигенов эритроцитов у людей с фе-
нотипом Rhnull. В основном это касается антигенов S, s, U, LW, Fy и i. Экспрессия антигенов S, s и, особенно, U на эритроцитах Rhnull существен-
но снижена. Как показали Schmidt и соавт. [598], почти половина сывороток
анти-S, анти-s и анти-U не реагировали с эритроцитами Rhnull S, Rhnull s и Rhnull U в антиглобулиновой пробе. В реакции солевой агглютинации эти образцы эри-
троцитов реагировали хорошо.
Таблица 4.25
Изменение экспрессии некоторых антигенов на эритроцитах Rh null
| Фенотип | Экспрессия антигенов | ||||||||
| S | s | U | LW | Fy5 | i | Rh29 total | D | ||
| Rhnull | ↓ | ↓ | ↓↓ | 0 | 0 | ↑ | 0 | 0 | |
| Rhmod | н | н | н | н | н | н | н | cледы | |
Примечание. ↓ – экспрессия снижена, ↑ – повышена, н – не изменена.
Race и Sanger [544] пришли к выводу, что связывание антител с указанными антигенами при фенотипе Rhnull слабее, чем в норме. Слабофиксированные ан-титела легко удаляются с поверхности эритроцитов в процессе их отмывания при постановке антиглобулиновой пробы. Этим и объясняются разные резуль-таты, получаемые в антиглобулиновой пробе и реакции солевой агглютинации.
Антигены LW a, LW b, LW ab и Fy5 в эритроцитах Rhnull отсутствуют, так же как и антигены Rh-Hr. Гены LW и Fy расположены на хромосоме 1 рядом
и локусом RН и, по-видимому, претерпевают те же изменения, что и гены ре-зус. Отсутствие указанных антигенов наблюдали независимо от регуляторного
(X O r / X O r) или аморфного (r=/r=) типа Rhnull.
Schmidt и соавт. [598] отметили повышенную экспрессию антигена i у жен-щины Rhnull, больной анемией. Sturgeon [638] констатировал некоторое повыше-ние экспрессии антигенов M и N при фенотипе Rhnull.
Race и Sanger [544] исследовали эритроциты 5 лиц Rhnull двумя сериями сыво-роток анти-En a и установили, что эритроциты Rhnull En(a + ) реагируют значительно
253
сильнее, чем эритроциты En(a + ) обычных людей. Указанные авторы отметили так-же повышение экспрессии антигенов Kidd и Dombrock на клетках Rhnull.
Природа дефицита некоторых антигенов на эритроцитах Rhnull понемногу проясняется. Установлено (Dahr и соавт. [241]), что эритроциты Rhnull обоих ге-нетических типов содержат 30–40 % гликофорина В (структура, которая несет на себе детерминанты S, s и U), т. е. в 2–3 раза меньше, чем эритроциты U + с нормальным фенотипом Rh. Дефицит антигенов LW на эритроцитах Rhnull обу-словлен отсутствием в их мембране соответствующего LW-гликопротеина (Bloy
В соавт. [173]).
Rh mod
У отличие от фенотипа Rhnull, при котором все антигены Rh отсутствуют, фе-нотип Rhmod (модифицированный) содержит небольшое, иногда исчезающее ко-личество Rh-антигенов. Некоторые образцы эритроцитов, идентифицирован-ные вначале как Rhnull, при более детальном исследовании оказывались Rhmod. Как показали Stevenson и соавт. [629], Rhnull – не всегда нуль.
Фенотип Rhmod появляется в результате воздействия на локус RH регулятор-ного (супрессорного) гена X Q. Этот ген, как полагают Chown и соавт. [225, 226], является аллелем гена X o r или самостоятельным геном, расположенным в ином, чем X o r, локусе, регулирующем количество Rh-субстанции.
Семейные исследования подтвердили, что лица с фенотипом Rhmod име-ют нормальные гены RH [225, 226]. Таким образом, генетическая основа Rhmod всегда регуляторного типа, в отличие от Rhnull, который может быть обусловлен аморфным типом наследования.
Количество антигенов Rh на эритроцитах лиц Rhmod значительно варьирует. В одних случаях Rh-антигены выявляют прямой реакцией агглютинации, в других – только с помощью высокочувствительного метода адсорбции – элюции. Mallory
и соавт. [Vox. Sang., 1976, V. 30, P. 430] нашли, что исследованные ими образцы эритроцитов Rhmod содержат только 1,2 % ожидаемого количества D-антигена.
Содержание гликофорина В на эритроцитах Rhmod уменьшено до 70 % от нормы.
Дата: 2019-02-24, просмотров: 385.