| Источник получения | Специфичность | |
| Dolichos biflorus | Семена | Анти-A1 |
| Phaseolus limensis | То же | Анти-А |
| Phaseolus lunatus | " | То же |
| Helix pomatia | Печень | " |
| Helix hortensis | То же | " |
| Fomes fometarius | Мицелий | Анти-В |
| Ptilota plumosa | Органеллы | То же |
| Salmo salar | Икра | " |
| Sophora japonica | Семена | Анти-АВ |
| Phlomis fructosa | То же | То же |
| Bandeiraea simplicifolia | " | " |
| Ulex europaeus | " | Анти-Н |
| Lotus tetragonolous | " | Анти-Н (HI) |
| Anguilla anguilla | Сыворотка | Анти-Н (HI) |
| Cystisus sessifolius | Семена | Анти-Н |
| Laburnum alpinum | То же | То же |
Антиген С
Существование антигена С в системе АВО можно считать доказанным. Он содер-жится в эритроцитах A(II), B(III) и AB(IV). В эритроцитах O(I) антиген С отсутствует.
Основанием для такого утверждения служат эксперименты Wiener и со-авт. [213, 224, 228–230], показавшие, что сыворотки Оαβ(I), помимо α- и β-агглютининов, реагирующих с эритроцитами А и В, содержат еще одну разно-видность естественных групповых антител – С-агглютинины (по Винеру анти-С-антитела), которые реагируют с эритроцитами А и В перекрестно [224, 229, 230].
80
Феномен перекрестного реагирования сывороток Оαβ(I) впервые описали Landsteiner и Witt в 1926 г. [134] как парадокс, нарушающий стройную концеп-цию системы АВО, постулирующую 2 агглютиногена – А и В, и два агглюти-нина – α и β. Вопреки ожиданиям исследователей элюаты, полученные при ад-сорбции сывороток Оαβ(I) эритроцитами А, реагировали не только с эритроци-тами А, но и с эритроцитами В. Такой же результат наблюдали после адсорб-ции сывороток Оαβ(I) эритроцитами В: элюат реагировал с эритроцитами В и А.
При адсорбции смеси сывороток Аβ(II) и Вα(III) перекрестного реагирования антител не происходило: элюат с эритроцитов А реагировал только с эритроцита-ми А, элюат с эритроцитов В – только с эритроцитами В (Wiener и соавт. [229]).
Адсорбционные пробы с сывороткой Оαβ(I), а именно возможность удалить с помощью эритроцитов А агглютинины β, а с помощью эритроцитов В – агглю-тинины α, указывают на то, что антигены А и В содержат общий компонент, ко-торый и является, по Винеру, антигеном С.
Весьма убедительными казались доводы оппонентов, отрицавших существо-вание антигена С. Так, Dodd, Lincoln, Boorman [91, 146], Bird [80] полагали, что анти-С-антитела представляют собой химически сшитые, несепарируемые, αβ-молекулы, способные реагировать с антигенами А и В. Таким образом, анти-ген С на эритроцитах А и В – это лишь видимость.
Другие видные исследователи (Race и Sanger [184], Доссе [25], П.Н. Косяков [35], Прокоп [56]) также указывали, что α- и β-агглютинины лиц группы О(I) не являются простой смесью. Логично допустить, что в процессе синтеза α-агглютининов и одно-временно β-агглютининов какая-то часть из них может быть собрана как αβ-антитела.
Важным аргументом против существования антигена С явился также тот факт, что до настоящего времени не найдены индивиды, в эритроцитах которых антиген С присутствует в чистом виде без антигенов А и В.
По мнению Wiener [229], некоторые образцы эритроцитов Аx (А4, А5) как раз и являются носителями антигена С в чистом виде. Эти эритроциты не содержат ан-тигена В, а количество антигена А в них крайне мало. Особенностью этих эри-троцитов является то, что они не реагируют с сыворотками Вα(III), но реагируют с сыворотками Оαβ(I), так как сыворотки Вα(III) содержат только α-агглютинины, а сыворотки Оαβ(I) наряду с α-агглютининами имеют анти-С-антитела. Более того, у лиц Аx в сыворотке крови иногда содержатся α1-экстраагглютинины, поэтому нельзя исключать, что антигены А4 и А5 таковыми на самом деле не являются, т. е. не относятся к группе А, а представляют собой антиген С.
Race и Sanger [184] не разделяли точку зрения Винера, указывая на то, что некоторые сыворотки Вα(III) слабо, но все-таки реагируют с эритроцитами А4, А5 [184]. Кроме того, в слюне людей А4, А5 находят растворимый антиген А, нейтрализующий α-агглютинины сывороток Вα(III), что свидетельствует о принадлежности антигенов А4 и А5 к группе А.
Дискуссия относительно существования антигена С в системе АВО, продол-жавшаяся до 1970-х годов, в последующие годы не возобновлялась в связи с
81
отсутствием новых данных, а также некоторым скептическим отношением им-муносерологов к этому вопросу, не представляющему, по их мнению, большого значения для практики переливания крови.
На наш взгляд, перекрестно реагирующие антитела имеют значение в транс-фузиологии. Их присутствие у реципиентов, по-видимому, усугубляет тяжесть посттрансфузионных осложнений. Об этом свидетельствует то обстоятельство, что среди реципиентов О(I), которым перелили кровь другой группы, осложне-ния протекают более тяжело и риск летального исхода выше, чем у реципиен-тов A(II) и B(III), которым перелили кровь другой группы. Однако соответству-ющие иммуносерологические исследования, подтверждающие или опроверга-ющие высказанное положение, не были проведены.
Вместе с тем в литературе накапливались сведения о том, что перекрестно ре-агирующие антитела, а стало быть и антиген С, имеют значение в акушерстве.
Rosenfield в 1953 г. [188, 189], исследуя групповые антитела у новорожден-ных и их матерей, обратил внимание на 2 обстоятельства: 1 – если мать и плод имели группу О(I), то у новорожденных α- и β-агглютинины в сыворотке крови присутствовали чаще, чем в случаях, когда мать и плод имели группу A(II) или оба относились к группе B(III); 2 – у женщин с О(I) группой крови ГБН разви-валась чаще, чем у рожениц, имевших другие группы крови.
Unger и Wiener [213], Wiener и Wexler [231], подтвердив данные Rosenfield, показали, что у новорожденных О(I) от матерей О(I) через месяц после рожде-ния перекрестно реагирующие αβ-антитела исчезали, что свидетельствовало об их материнском происхождении.
Kochwa и Rosenfield установили, что перекрестно реагирующие антитела анти-А,В относятся к глобулинам 7Sγ2, которые легко проникают через плацен-ту, поскольку размер их молекул меньше, чем у других антител.
Разделяя взгляды Винера, мы предприняли попытку найти подтверждение его концепции, а именно того, что перекрестные реакции в системе АВО об-условлены антигеном С эритроцитов, а не комбинированными αβ-антителами.
Для начала представлялось целесообразным получить собственные данные о ха-рактере перекрестных реакций, свойственных сывороткам Оαβ(I) [20, 21, 23, 24, 47].
Материал и методы. Исследовали сыворотки крови 100 доноров, имеющих груп-пу крови О(I), мужчин и женщин. Каждую сыворотку титровали с эритроцитами А и В до и после адсорбции эритроцитами А, В и О. Титр сывороток устанавливали кратным их разведения 0,9 % NaCl. В качестве контроля первые 3 разведения сыворотки тести-ровали эритроцитами О(I). Адсорбцию антител проводили при комнатной температуре 20–22 оС в течение 1 ч в соотношении 2 объема сыворотки + 1 объем плотно упакован-ного гомогенизированного осадка трижды отмытых эритроцитов. Устойчивость анти-тел к унитиолу (2,3-димеркаптопропансульфонат натрия – CH2SH-CH-SH-CH2SO3Na) оценивали титрованием сыворотки после добавления равного объема 1,25 % раствора этого реактива и инкубации полученной смеси в течение 2 ч при температуре 37 оС. Температурную устойчивость антител исследовали посредством прогревания сыворо-ток при температуре 56 и 70 оС в течение 1 ч и 10 мин соответственно.
82
Из 100 исследованных сывороток группы О(I) 52 не содержали перекрестно реагирующих антител. При уменьшении титра α-антител на 4–8 ступеней после адсорбции этих сывороток эритроцитами А β-антитела оставались в том же ти-тре, что и до адсорбции. Аналогичный результат наблюдали после адсорбции этих сывороток эритроцитами В. Титр β-антител снижался на 4–8 ступеней, в то время как титр α-антител оставался прежним (табл. 3.7).
Таким образом, 52 % сывороток Оαβ(I) не обладали способностью к пере-крестному реагированию, 48 % сывороток содержали перекрестно реагирую-щие антитела. Перекрестное реагирование было симметричным и асимметрич-ным, в частности, 20 из 48 сывороток показали симметричное перекрестное ре-агирование, т. е. независимо от того, какими эритроцитами (А или В) проводи-ли их адсорбцию, титр α- и β-агглютининов снижался пропорционально.
Другие 28 сывороток показали асимметричное перекрестное реагирование. В 15 сыворотках после адсорбции эритроцитами А снижался титр обоих агглютининов (α
и β), а после адсорбции эритроцитами В убывал титр только β-агглютининов, титр α-антител оставался прежним. В 13 сыворотках, наоборот, снижение титра обоих аг-глютининов наблюдали после адсорбции эритроцитами В. Адсорбция эритроцитами А снижала титр α-агглютининов, но не влияла на титр β-антител. Таким образом, пе-рекрестное реагирование по своему характеру может быть симметричным, асимме-тричным в сторону А-антигена и асимметричным в сторону В-антигена.
| Таблица 3.7 | ||||||||||||||||
| Варианты перекрестной адсорбции сывороток О αβ (I) | ||||||||||||||||
| Категория | Серия | Титр | Титр после адсорбции эритроцитами | |||||||||||||
| до адсорбции | A(II) | B(III) | О(I) | |||||||||||||
| сыворотки | сыворотки | с эритроцитами | с эритроцитами | с эритроцитами | с эритроцитами | |||||||||||
| A | В | О | А | В | О | А | В | О | А | В | О | |||||
| 399 | 1 : 256 | 1 : 256 | 0 | 1 : 2 | 1 : 256 | 0 | 1 : 256 | 0 | 0 | 1 : 2 56 | 1 : 256 | 0 | ||||
| I | 626 | 1 : 32 | 1 : 8 | 0 | 0 | 1 : 8 | 0 | 1 : 32 | 0 | 0 | 1 : 32 | 1 : 8 | 0 | |||
| 997 | 1 : 256 | 1 : 128 | 0 | 0 | 1 : 128 | 0 | 1 : 256 | 0 | 0 | 1 : 256 | 1 : 128 | 0 | ||||
| 846 | 1 : 64 | 1 : 32 | 0 | 0 | 1 : 32 | 0 | 1 : 64 | 0 | 0 | 1 : 64 | 1 : 32 | 0 | ||||
| 129 | 1 : 128 | 1 : 128 | 0 | 1:2 | 1 : 32 | 0 | 1 : 64 | 0 | 0 | 1 : 128 | 1 : 128 | 0 | ||||
| II | 272 | 1 : 256 | 1 : 128 | 0 | 0 | 1 : 64 | 0 | 1 : 64 | 0 | 0 | 1 : 256 | 1 : 128 | 0 | |||
| 397 | 1 : 256 | 1 : 64 | 0 | 0 | 1 : 32 | 0 | 1 : 32 | 1 : 2 | 0 | 1 : 256 | 1 : 64 | 0 | ||||
| 488 | 1 : 256 | 1 : 32 | 0 | 0 | 1 : 8 | 0 | 1 : 32 | 0 | 0 | 1 : 256 | 1 : 32 | 0 | ||||
| 127 | 1: 256 | 1 : 128 | 0 | 1 : 2 | 1 : 64 | 0 | 1 : 256 | 1 : 2 | 0 | 1 : 256 | 1 : 128 | 0 | ||||
| III | 855 | 1 : 128 | 1 : 64 | 0 | 0 | 1 : 32 | 0 | 1 : 128 | 0 | 0 | 1 : 128 | 1 : 64 | 0 | |||
| 505 | 1 : 256 | 1 : 128 | 0 | 0 | 1 : 64 | 0 | 1 : 128 | 0 | 0 | 1 : 256 | 1 : 128 | 0 | ||||
| 705 | 1 : 512 | 1 : 256 | 0 | 0 | 1 : 128 | 0 | 1 : 512 | 0 | 0 | 1 : 512 | 1 : 256 | 0 | ||||
| 400 | 1 : 128 | 1 : 128 | 0 | 0 | 1 : 128 | 0 | 1 : 64 | 0 | 0 | 1 : 128 | 1 : 128 | 0 | ||||
| IV | 424 | 1 : 256 | 1 : 64 | 0 | 0 | 1 : 64 | 0 | 1 : 128 | 0 | 0 | 1 : 256 | 1 : 64 | 0 | |||
| 454 | 1 : 256 | 1 : 128 | 0 | 1 : 2 | 1 : 128 | 0 | 1 : 128 | 1 : 2 | 0 | 1 : 256 | 1 : 128 | 0 | ||||
| 351 | 1 : 256 | 1 : 128 | 0 | 0 | 1 : 128 | 0 | 1 : 128 | 0 | 0 | 1 : 256 | 1 : 128 | 0 | ||||
Примечание. Категория I – сыворотки, не содержащие перекрестно реагирующих антител,
II – сыворотки, содержащие антитела, дающие симметричные перекрестные реакции, III и IV – сыворотки, содержащие антитела, дающие асимметричные перекрестные реакции.
83
На наш взгляд, перекрестное реагирование обусловлено не αβ-агглютининами, как полагала Dodd и соавт., а антителами αС, βС, что в большей мере согласуется
с концепцией Винера, а также полученными нами данными. На рис. 3.2 представ-лена предполагаемая модель этих антител.
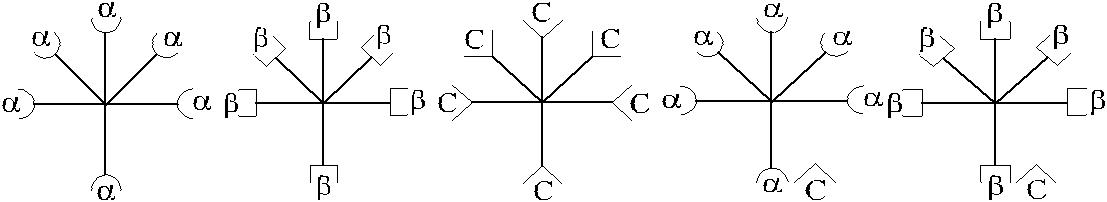
| 1 | 2 | 3 | 4 | 5 |
| – антиген А по Винеру | – антиген В по Винеру |
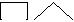
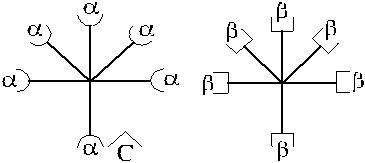
Рис. 3.2. Возможные варианты изогемагглютининов в сыворотке Оαβ(I).
1 – α-изогемагглютинины; 2 – β-изогемагглютинины, 3 – С-изогемагглютинины;
4 – αС-изогемагглютинины; 5 – βС-изогемагглютинины.
Представленная структурная модель изогемагглютининов позволяет объ-яснить характер перекрестного реагирования сывороток Оαβ(I). Сыворотки, не вызывающие перекрестных реакций, содержат только α- и β-антитела (рис. 3.3, п. 1). Сыворотки, вызывающие симметричные перекрестные реакции, содержат, помимо α- и β-антител, антитела αС и βС или анти-С-антитела (рис. 3.3, п. 4). Сыворотки, вызывающие асимметричные перекрестные реакции, содержат α, β и одну из разновидностей антител – αС или βС (рис. 3.3, п. 2, 3).
1. Перекрестная реакция отсутствует 2. Асимметричная перекрестная реакция
(α-агглютинины не могут адсорбироваться на (α-агглютинины могут адсорбироваться на
эритроцитах В, β-агглютинины не могут адсор- эритроцитах В за счет присутствующего на них
бироваться на эритроцитах А). антигена С).
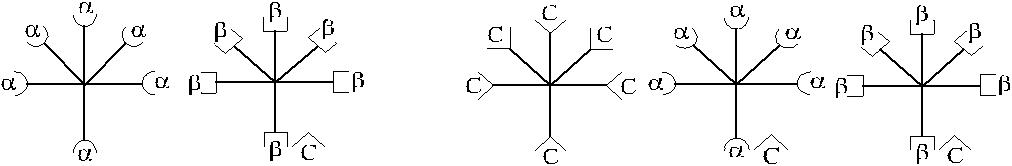
3. Асимметричная перекрестная реакция 4. Симметричная перекрестная реакция
(β-агглютинины могут адсорбироваться на эри- (1-й вариант: β-агглютинины адсорбируются на
троцитах А за счет присутствующего на них ан- эритроцитах А, α-агглютинины адсорбируются
тигена С). на эритроцитах В за счет рецептора анти-С, 2-й
вариант: анти-С-агглютинины адсорбируются
на эритроцитах А и В).
Рис. 3.3. Комбинации агглютининов, обусловливающие различные варианты пере-крестного реагирования.
84
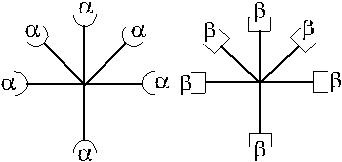
Рассмотрим модель асимметричного реагирования сыворотки, которая со-держит агглютинины в сочетании, представленном на рис. 3.4.
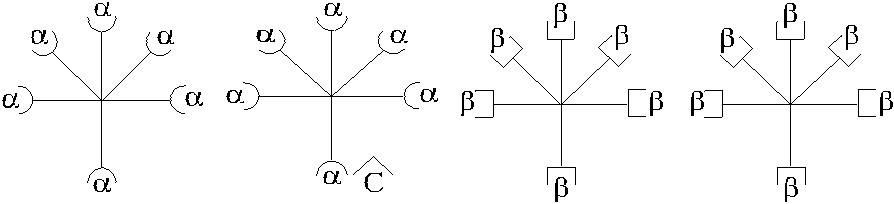
Рис. 3.4. Вариант асимметрично реагирующей сыворотки.
Согласно схеме сыворотка содержит 50 % α-агглютининов, 75 % – β. Валентность анти-С реагирует как α и β. При адсорбции такой сыворотки эритро-цитами А α-агглютинины будут удалены полностью. Иными словами, из сыворот-ки будет удалено 50 % антител. При адсорбции сыворотки эритроцитами В будет удалено 75 % антител (50 % β + 25 % α) за счет рецептора анти-С. В случае, если рецептор анти-С связан не с α-, а с β-агглютининами, асимметричное реагирова-ние проявят эритроциты А, унося на своей поверхности 25 % β-агглютининов.
Как далее нами показано, перекрестно реагирующие антитела так же, как иммун-ные антитела АВО и Rh, относятся к классу IgG, устойчивы к унитиолу, лучше реаги-руют с отмытыми, чем неотмытыми эритроцитами, в отличие от иммунных (термо-стабильных) антител являются термолабильными (утрачивают активность после про-гревания сыворотки при 70 оС в течение 10 мин). В противоположность классиче-ским иммунным антителам IgG, которые адсорбируются на эритроцитах, но не вызы-вают их агглютинации, перекрестно реагирующие антитела непосредственно агглю-тинируют эритроциты в солевой среде. В низкой концентрации они утрачивают спо-собность агглютинировать эритроциты, однако сенсибилизируют их подобно непол-ным антителам и хорошо выявляются в непрямой антиглобулиновой пробе Кумбса.
Сыворотки лиц Оαβ(I), очевидно, содержат несколько типов групповых анти-тел: изогемагглютинины α и β IgM, α и β IgG, αС и βС IgG, анти-С IgG, иммунные
α и β IgM, иммунные α и β IgG, что закономерно обусловливает перекрестные ре-акции с различными групповыми антигенами.
Важным аргументом, подтверждающим концепцию Винера о существовании антигена С, явились эксперименты по получению специфических антисыворотк. В частности, при иммунизации мышей эритроцитами А(II) и В(III) нами [21] были получены моноклональные антитела со специфичностью анти-АВ (МКА-АВ).
Материал и методы. Трехмесячным самкам мышей BALB / с вводили по 0,1 мл от-мытых 0,9 % NaCl эритроцитов А(II) или В(III) внутрибрюшинно дважды с интерва-лом в 7 дней. Третью инъекцию производили внутривенно за 5 дней до слияния спле-ноцитов с клетками миеломы NS-1. Слияние осуществляли по стандартной методике
с использованием 45 % полиэтиленгликоля-1500 «Loba» и 5 % диметилсульфоксида. Эффективность гибридизации составляла 10–5. Клонировали гибридные клетки в сре-де НАТ без фидера. Антителопродуцирующие клоны отбирали с помощью реакции аг-глютинации в 96-луночных планшетах с отмытыми эритроцитами О(I), А(II) и В(III).
Класс моноклональных антител устанавливали по флюоресценции сенсибилизи-
85
рованных эритроцитов со специфическими антимышиными сыворотками, меченны-ми флюоресцеинизотиоцианатом «Sigma». Реакцию учитывали на флюоресцентном микроскопе «Opton». Иммуносерологические параметры моноклональных антител определяли с помощью общепринятых серологических методов: реакции агглютина-ции в солевой и коллоидной среде, непрямой пробы Кумбса.
Образцы моноклональных антител титровали до и после адсорбции эритроци-тами А(II), В(III) и О(I).
Адсорбцию антител проводили при комнатной температуре 20–22 оС в течение 1 часа в соотношении 2 объема сыворотки + 1 объем плотно упакованного гомогени-зированного осадка трижды отмытых эритроцитов.
Элюцию антител выполняли по методике, описанной Wiener [232], Feng и соавт. [102]: осадок трижды отмытых сенсибилизированных эритроцитов замораживали, за-тем оттаивали и добавляли к нему 50 % этиловый спирт в соотношении 1 : 9. Смесь вновь замораживали, оттаивали и отмывали дистиллированной водой. К полученно-му осадку стромы сенсибилизированных эритроцитов добавляли равный объем 0,15 М NaCl и инкубировали при температуре 37 оС 1 ч, далее смесь центрифугировали, полу-ченную надосадочную жидкость (элюат) испытывали на специфичность и активность.
Устойчивость антител к унитиолу (2,3 димеркаптопропансульфонат натрия – CH2SH-CH-SH-CH2SO3Na) оценивали посредством титрования сыворотки после до-бавления равного объема 1,25 % раствора этого реактива и инкубации полученной смеси в течение 2 ч при температуре 37 оС.
Нейтрализацию антител производили водорастворимыми группоспецифическими субстанциями, фруглюмином А и В, выделенным из желудка свиней и лошадей (про-изводства Белорусского НИИГПК), из расчета 1 мг сухого фруглюмина на 1 мл МКА.
Из полученных гибридом отобрали 8 продуцирующих МКА анти-АВ. Серии 1–7 МКА анти-АВ получили при иммунизации эритроцитами А(II), серию 8 МКА анти-АВ – эритроцитами B(III). Все серии антител агглютинировали эритроциты А1(II), А2(II) и В(III) и не реагировали с эритроцитами О(I). Титр МКА анти-АВ с эритроци-тами А1 соответствовал 1 : 16–1 :1 28, с эритроцитами B(III) – 1 : 8–1 : 64 (табл. 3.8).
Таблица 3.8
Дата: 2019-02-24, просмотров: 467.