Организма
Гормоны — это органические сигнальные молекулы беспроводникового системного действия. Они распознаются рецепторами и влияют на экспрессию генов и активность ферментов в клетках-мишенях на удалении от места своей продукции. Гормоны не являются ни катализаторами, как энзимы, ни кофермента-ми, как витамины, ни пластико-энергетическим сырьем, как нутриенты.
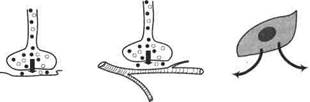
Э. Г. Старлинг образно называл гормоны вестни ками. Их роль — информационная, поскольку в зависимости от своего взаимодействия с рецепторами и от состояния клеток-мишеней в контексте других сигнальных эффектов они включают и выключают на генетическом уровне определенные клеточные программы или модулируют эпигенетически эффективность их осуществления. Нейротрансмиттеры (например, аце-тилхолин) в отличие от гормонов действуют только в пределах синапсов, куда попадают по проводникам, а аутокоиды (например, производные арахидоновой кислоты) действуют зонально и инактивируются в очаге своей продукции (рис. 1).
Приведённые градации относительны, так как одна и та же молекула в разных ситуациях может выступать в любой из этих трёх ипостасей, примером чего служат катехоламины, в частности, норадреналин. Нередко гормоны распространяются лишь в пределах ограниченной циркуляторной сети. Так происходит с гипоталами-ческими либеринами и статинами, а также с многими гормонами желудочно-кишечного тракта, которые предохраняются от системного распространения, так как инкретируются в портальные системы, снабжённые собирающими сосудами между двумя капиллярными сетями. Такой гормон попадает в кровь в пределах первой капиллярной сети (например, в гипоталамусе, клубочках почек или непарных органах брюшной полости) и с высоким сродством связывается рецепторами клеток, снабжаемых вторичной капиллярной сетью (например, в гипофизе, в районе петли Генле в почках или в печени).
В отношении обмена веществ, клеточной пролиферации, роста тканей, их дифференцировки, созревания и долговременных адаптивных реакций гормоны служат основными регуляторами, определяющими направленность и эффективность этих процессов. Нейротрансмиттеры в своих воздействиях на эти стороны жизнедеятельности или зависят от регулирующих эффектов гормонов, или даже опосредуют свои влияния через гормоносекретирующие клетки. Ценными особенностями гормональной регуляции являются ее беспороговый принцип и большое разнообразие пермис-сивных взаимодействий гормонов, исключающее господство стереотипии.
| Эндокринный |
| Паракринный |
|
|
| Нейромедиаторный Нейроэндокринный Панокринный |
В то же время проводниковая нервная регуляция, осуществляемая на основе пороговых стереотипных отве-
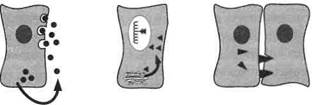
Наружный Внутренний Юкстакринный
аутокринный аутокринныи
Рис. 1. Различные типы химических регуляторных взаимодействий между клетками (по Дж. Докрею, К. Р. Хопкинсу, 1982)
тов, играет приоритетную роль в управлении движением, в сборе афферентной информации, а также в регуляции быстрых типовых реакций на различные стимулы.
Классическим примером ключевой роли гормонов как регуляторов онтогенеза служит наступление метаморфоза у головастиков под влиянием тироксина и трийодтиронина.
Гормональная, нервная, а также иммунная регуляции клеточных функций неразрывно связаны между собой (Зайчик А.Ш., Чурилов Л.П., 1999, 2001). Примерами могут служить необратимое нарушение развития центральной нервной системы при врождённом гипоти-розе* {болезнь Фаггё), а также наличие секреторной иннервации у многих эндокринных желёз (эпифиза, мозгового вещества надпочечников, щитовидной железы, островков Лангерганса). В XVIII столетии далеко опередивший своё время французский врач Теофиль де Бордю (1775) высказал гениальную догадку о том, что все органы посылают в кровь свои специфические хи мические сигналы, управляя жизнедеятельностью тела. Он писал: «Каждый орган служит фабрикой и лабо раторией специфического гуморального агента, который по приготовлении и приобретении индивидуально присущих ему свойств возвращается в кровь. Кровь обладает специфическими свойствами, приобретенными в органах, через которые она проходит. Каждый орган посылает в нее свою эманацию... Таким образом, кровь несет в своем потоке экстракты всех органов, необходимые для жизни целостного организма...».
К 1844 году развитие анатомии обеспечило И. Мюллера достаточным количеством фактов, чтобы сделать обобщение о делении желез на органы, занятые внешней и внутренней секрецией. Последние, по мнению этого немецкого учёного, всегда лишены выводных протоков и посылают секрет в кровь и/или в лимфу. В разряд их, по Й. Мюллеру, попали не только щитовидная железа или надпочечники, но и селезёнка, лимфоузлы, печень и т.д. Характерно, что классики той эпохи не выделяли какие-то особые молекулы исключительно как сигналы, а рассматривали любое химическое вещество, которым «железа» обогащает кровь, как «инкрет». Например, К. Бернар (1857) характеризовал глюкозу как инкрет печени.
Первым химически индентифицированным гормоном был адреналин мозгового вещества надпочечников (Такамине Ж., 1901). Само понятие «гормон» не-
* С 1985 г. Международным конгрессом эндокринологов рекомендовано применять медицинские термины с корнем «тиро» вместо ранее употреблявшегося «тирео».
10
многим позже ввел первооткрыватель секретина Э.Г. Старлинг (1905), определив его как «вещество, вырабатываемое в одной части организма и переносимое кровью или лимфой к какой-либо другой части, функция которой вследствие этого изменяется».
Чуть позже Н. Ленде (1909) ввел термины «эндок ринология» и «эндокринные железы», и концепция гормональной регуляции после более чем 230-летнего существования добилась, наконец, полноправного положения в клинической и экспериментальной медицине.
Строго говоря, генетические программы гормоно-образования имеются в каждой клетке. Развитие в XX веке точных и чувствительных иммуноконкурентных методов определения содержания гормонов (радиоиммунологического, а позже — иммуноферментного) позволило доказать, что гормоны не обязательно являются продуктами анатомически обособленных эндокринных желёз. Эндокринная железа может представлять собой отдельный орган (например, щитовидная железа) или часть более сложного органа (например, островки Лангерганса поджелудочной железы).
Более того, хотя выделение гормонов системного или дистантного действия в кровь и лимфу (эндокринная сигнализация) присуще не каждой клетке, но оно и не сосредоточено только лишь в особых образованиях, как полагал Й. Мюллер. Гормон может быть продуктом расположенных в разных органах и тканях групп клеток и отдельных клеток так называемой диф фузной (дисперсной) эндокринной системы.
Ещё в 1938 г. гистолог Ф. Фейртер сформулировал понятие о диффузных эндокринных клетках желудочно-кишечного тракта. Подобные клетки были обнаружены позже и во многих других органах.
Обобщая подобные данные, А. Дж. Э. Пирс (1968) выдвинул концепцию АПУД-системы и назвал подобные диффузные эндокриноциты «апудоцитами» (от англ. APUD — amine precursor uptake and decarboxyla tion —захват и декарбоксилирование аминных предшественников). Он имел в виду, что эти клетки способны к синтезу гормонов — производных аминокислот. Большая часть клеток АПУД-системы * * происходит из нервного гребня. Многие клетки энтодермального и мезен-химального происхождения могут под влиянием внешних стимулов приобретать свойства АПУД-клеток. Все эти клетки способны накапливать триптофан, гистидин и тирозин и превращать их путем декарбоксилирования в медиаторы {серотонин, дофамин и гистамин). Впослед-
*Современный, более широкий по значению термин — диффузная нейроэндокринная система (ДНЭС — DNES).

 ствии оказалось, что им присуща и продукция аминов, и выработка регуляторных пептидов, причём часто (вопреки догме «одна клетка -— один гормон») — в пределах одного клеточного типа. Так, В-клетки островков Лангерганса выделяют и инсулин, и ГАМК. Все эти клетки, имеющие черты как эндокриноцитов, так и параней-ронов, обладают маркерами последних: хромогранином А, ферментами декарбоксилазой и нейрон-специфической енолазой и, по существу, являются нейроэндокрин-ными (Малашхия Ю.А. с соавт., 1990). Первоначально А. Дж. Э. Пирс постулировал, что все они расселяются по организму из нейроэктодермы нервного гребешка, но позже А. Эндрю (1981) доказала, что диффузные эндокриноциты могут иметь также и энто-дермальное, и мезодермальное происхождение.
ствии оказалось, что им присуща и продукция аминов, и выработка регуляторных пептидов, причём часто (вопреки догме «одна клетка -— один гормон») — в пределах одного клеточного типа. Так, В-клетки островков Лангерганса выделяют и инсулин, и ГАМК. Все эти клетки, имеющие черты как эндокриноцитов, так и параней-ронов, обладают маркерами последних: хромогранином А, ферментами декарбоксилазой и нейрон-специфической енолазой и, по существу, являются нейроэндокрин-ными (Малашхия Ю.А. с соавт., 1990). Первоначально А. Дж. Э. Пирс постулировал, что все они расселяются по организму из нейроэктодермы нервного гребешка, но позже А. Эндрю (1981) доказала, что диффузные эндокриноциты могут иметь также и энто-дермальное, и мезодермальное происхождение.
Еще в 1928 г. М. Шеррер открыл явление нейро-секреции и описал гормональную активность нейронов гипоталамуса. Таким образом, им были заложены представления о нейросекреции. Принципиальное значение имело неожиданное наблюдение, что практически все пептидные гормоны, первоначально найденные в желудочно-кишечном тракте (гастрин, вещество Р, холецистокинин, вазоактивный инте-стинальный полипептид, инсулин, глюкагон и другие) содержатся также и в центральной нервной системе. И, напротив, некоторые, первично открытые в ней нейропептиды (соматостатин, нейротензин, эндогенные опиаты и пр.), позже были найдены как инкреторные продукты диффузных эндокриноцитов кишечника и островков Лангерганса. Более того, выяснилось, что эти пептиды, будучи гормонами в желудочно-кишечном тракте, служат нейротрансмитте-рами в пептидэргических системах мозга или выделяются в безымпульсном режиме после антидромного распространения по афферентным нервным проводникам теми же нейронами, которые оперируют и аминными нейромедиаторами (Докрей Г. Дж., 1978; Поляк Дж. 1 YL , Блум С. М., 1977).
В настоящее время считают, что одни и те же сигнальные молекулы служат в мозге глобальным средством химического кодирования тех форм нейронной активности, которые связаны с отдельными древнейшими функциями (половым и пищевым поведением, поддержанием водно-электролитного баланса, позитивным и негативным подкреплением, памятью), а в других органах (например, в желудочно-кишечном тракте) используются локально как паракринные аутокоиды или гормоны. Более того, по У. Кэннону и А. Розенблюту (1951), антидромное распространение и секреция афферентными нервами таких нейропептидных гормонов служит реальной основой явления «нервной трофики». В насто-
ящее время развитие учения о нейропептидах привело к демистификации понятия «нервная трофика», которое трактуется, по существу, как эндокринная функция ней-роцитов. А. Дж. Э. Пирс (1977) прямо заключил, что «апудоциты продуцируют пептиды, действующие и как гормоны, и как нейромедиаторы. Они действуют как эффекторы третьего звена, поддерживая или модулируя функции соматических и вегетативных нейронов, служа фактически трофическими субстанциями как для нейронов, так и для соматических клеток». Вышеизложенные факты привели к освоению эндокринологами концепции о нейроэндок- ринном взаимодействии.
С открытием дисперсной эндокринной системы предметными стали вопросы эндокринной функции жировой ткани (лептин), сердца (атриопептины), почек (ренин, эритропоэтин, производные кальциферола), печени (ангиотензиноген, производные кальциферола) и других органов. Оказалось, что многие диффузные эндокриноциты выделяют прогормоны-предше-ственники, а активные гормоны совсем не обязательно должны вырабатываться в конкретной железе, но мо гут даже формироваться вне клеток — в крови. Примером служат ангиотензины II и III , предшественник которых (ангиотензин I ) возникает в печени и в диффузных эндокриноцитах разной локализации, а активные гормоны образуются прямо в плазме за счёт про-теолитического эффекта почечного ренина и лёгочной ангиотензин-конвертазы.
Наконец-то, можно сказать, разрушилась та искусственная «берлинская стена», которая делила непримиримых представителей лагеря нервистов и лагеря гу-моралистов, сломавших немало копий в попытке выяснить «кто главнее в организме?». Мозг фактически оказался крупнейшей эндокринной железой, а обе формы регуляции — двумя сторонами нейроэндокринной сети, в которой их функции не конкурируют, но разумно делят сферы преимущественной применимости и постоянно взаимодействуют.
Дальнейшее развитие эндокринологии поставило вопрос о том, что и иммунная система, функции которой также основаны на комплементарном взаимодействии химических сигналов и рецепторов, не остаётся в стороне от нейроэндокринного аппарата. Более того, в настоящее время в эндокринологии принято исходить из существования иммунонейроэндокринных взаимодействий и триединого коммуникативно-регуляторного интегрирующего аппарата (КРИА), включающего не только нейроэндокринную, но и иммунную системы.
Еще в 60-е годы XX столетия была раскрыта эндокринная роль тимуса. Эта уникальная эндокрин-
11
ная железа не только посылает в системный кровоток типичные гормоны (тимозин и тимулин), но и служит местом паломничества лимфоидных клеток-мишеней, которые, проходя через нее, подвергаются воздействию множества местных паракринных и юк-стакринных (действующих в пределах межклеточного контакта) гуморальных регуляторов-цитокинов, определяющих пути их дифференцировки (Миллер Дж., 1961). Затем стало ясно, что цитокины лимфоидных и макрофагальных клеток способны изменять функции гипоталамуса и гипофиза, а также обладают гормоноподобным действием на периферические эндокринные железы. Более того, оказалось, что гормоны (особенно пептидные), а также и их рецепторы часто служат обьектом физиологического аутоиммунного ответа, а при патологии — мишенью аутоал-лергии. Из-за химического сходства (гомологии) многих биорегуляторов и рецепторов бактерий и высших организмов подобная аутоаллергия, имеющая мишенью эндокринную систему, бывает частой причиной первичных и вторичных, железистых и внежелезистых эндокринопатий. Аутоаллергия оказалась одним из самых частых и важных патогенетических механизмов эндокринных болезней. Так, например, рецептор микроорганизма Yersinia enterocolytica, подобный ти-ротропиновому рецептору фолликулярных клеток щитовидной железы, вызывает у индивидов, предрасположенных к аутоаллергии, перекрёстный аутоиммунный ответ, поражающий тироциты и приводящий, по современным представлениям, к диффузному токсическому зобу — болезни Базедова (Харис-сон Л., 1985).
Было показано существование в норме, при патологии и в экспериментальных условиях антиидиоти-пических антител, полностью или частично имитирующих либо блокирующих действие пептидных и аминокислотных гормонов на клетки-мишени. Все эти факты убедили современных эндокринологов в том, что иммунная система, биорегуляторными сигналами которой служат цитокины и антитела, в норме и при патологии включается в нейроэндокринные взаимодействия, вследствие чего и сложилась концепция им- мунонейроэндокринного коммуникативно-регу-ляторного интегрирующего аппарата — КРИА (Корнева Е.А., 1987; Зайчик А.Ш. с соавт., 1982, 1988, 2002; Беседовский X., 1989). Появилась даже оригинальная точка зрения, что некоторые лимфоциты представляют собой своего рода циркулирующую разновидность апудоцитов — теория нейроиммун-ного дифферона (Абрамов В.В., 1986). Аксиомой физиологии стало учение о влиянии центральной не-
рвной системы, в частности, гипоталамуса на иммунный ответ через вегетативные нервы и гормоны (Кор нева Е.А., 1962, 2003). Подробнее эти и другие вопросы иммунонейроэндокринологии освещены в ряде недавно изданных книг и обзоров (Зайчик А.Ш. с соавт., 1988; Зайчик А.Ш., Чурилов Л.П., 1999, 2001, 2002; Корнева Е.А., 2003).
Для целей данного руководства важно подчеркнуть, что современная эндокринология —это наука об эн докринной регуляции и ее нарушениях (а не только о функциях и болезнях эндокринных желез).
Метаболизм гормонов
В настоящее время открыто около 100 гормонов. Химически они подразделяются на три группы:
— пептиды и гликопротеиды;
— производные аминокислот;
— дериваты холестерина.
Важно отметить, что одна и та же клетка может вырабатывать одновременно или на разных стадиях своего онтогенеза разные гормоны одной химической группы, а иногда — даже гормоны разных химических групп. Например, адренокортикоциты в онтогенезе мигрируют от капсулы внутрь надпочечников, последовательно выделяя минералокортикоиды, глюко-кортикоиды и андрогены, а некоторые из них, вероятно, способны трансформироваться впоследствии в смешанные клетки, секретирующие и катехолами-ны.
БИОСИНТЕЗ ГОРМОНОВ
По современным данным, биосинтез гормонов протекает по-разному в зависимости от их химической природы и характеризуется следующими особенностями.
Пептидные гормоны
Пептидные гормоны образуются по схеме «пре-прогормон — прогормон — активный гормон», иллюстрируемой на рис. 2.
Вначале формируется гетерогенная ядерная рибонуклеиновая кислота, содержащая транскрипты экзонов и интронов, затем при сплайсинге интроновые копии удаляются, а из экзоновых формируется м-РНК препро-гормона, снабжённая полиадениловым хвостом, длина которого определяет срок ее жизни в клетке. На её основе путём трансляции возникает препрогормон, имеющий сигнальную универсальную последовательность нa N-конце, необходимую для переноса молекулы

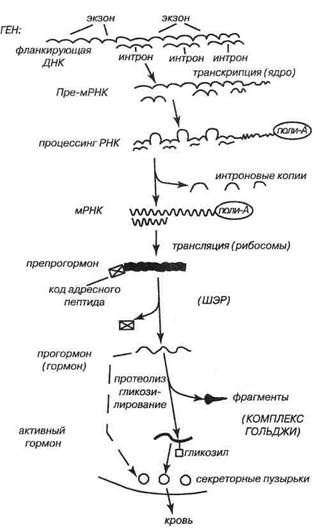
Рис. 2. Общая схема образования полипептидных гормонов
внутрь шероховатогозндоплазматического ретикулума (ШЭР).
Далее в комплексе Гольджи эндокриноцита происходит протеолитическое удаление (вырезывание) этого сигнального пептида, как правило, по связям между арги-ниновыми или аргининовым и лизиновым остатками. Образуется прогормон (например, проинсулин или прокалъцитонин), а иногда — сразу гормон (например, пролактин или гормон роста). Прогормон пакуется в секреторные пузырьки и затем непосредственно в них или в пределах эндоплазматического ретикулума происходит дальнейший протеолиз с образованием актив ного гормона.
Так осуществляется, например, вырезывание С-пептида с формированием инсулина. Впрочем, часто прогормон секретируется как таковой вместе с готовым гормоном, а протеолиз и вычленение актив ного гормона (гормонов) идёт частично на перифе-
рии, причем — по-разному в различных тканях или на разных стадиях гормонального ответа. Примером может служить процессинг гипофизарного проопиомела- нокортина (ПОМК) в надпочечниках при стрессе. В начале стресса ПОМК членится клетками гипофиза и надпочечников до АКТГ, а в конце начинает преобладать его протеолиз до опиатных пептидов. На разных стадиях онтогенеза один и тот же предшественник может процессироваться по-разному. Это относится, в частности, к ПОМК, который у плода и у взрослого образует по преимуществу различные варианты МСГ.
Некоторые пептидные гормоны (ТТГи все гонадо- тропины) перед секрецией гликозилируются в аппарате Гольджи, что способствует удлинению времени их жизни в крови.
Пептидные гормоны, как содержащие одну, так и имеющие две субъединицы, могут кодироваться одним геном {инсулин, паратгормон) или двумя генами (хорионический гонадотропин, лютеинизиру-ющий гормон). Иногда один гормон в разных органах может получаться из различных предшественников, закодированных неидентичными генами (соматос-татин). Часто, наоборот, один прогормон служит предшественником нескольких гормонов, получаемых из него путем посттрансляционного протеолиза. Например, проопиомеланокортин аденогипофиза служит сырьем для продукции АКТГ, в-липотропи-на, а-МСГ, в-МСГ, а-в-у-эндорфинов, энкефалина и кортикотропиноподобного промежуточного полипептида.
После транскрипции в разных эндокриноцитах одних и тех же генов гормонов возможен альтернативный сплайсинг гетерогенной ядерной РНК с образованием различных м-РНК, которые в дальнейшем приводят к трансляции неидентичных гормонов в различных тканях. Так, в С-клетках щитовидной железы формируется преимущественно калъцитонин, а в центральной нервной системе — пептид, связанный с геном калъ- цитонина, причём в обоих случаях — на основе общего транскрипта препрокалъцитонина.
Важной особенностью характеризуется синтез пептидных либеринов гипоталамуса. Показано, что образование многих из них (тиролиберина, гонадо-либерина, соматолиберина и пролактин-рилизинг фактора) идёт в цитозоле нейросекреторных клеток не рибосомальным путём. Оно не чувствительно к пу-ромицину и РНК-азе и напоминает механизм формирования глутатиона и антибиотических пептидов у микроорганизмов. Для осуществления этих процессов служит набор АТФ-зависимых цитоплазматических
13
ГЛЮКОКОРТИКОИДЫ
либерин-синтетаз (Гэррин Л. с соавт., 1971). Это ускоряет синтез либеринов и делает ответ ней-роэндокринных клеток очень лабильным и пластичным.
Стероидные гормоны
Стероидные гормоны синтезируются по совершенно иному пути. Сырьём служит либо холестерин (в коре надпочечников и гонадах — при образовании минера- локортикоидов, глюкокортикоидов, прогестинов, андрогенов и эстрогенов), либо 1 -дегидрохолесте-
МИНЕРАЛОКОРТИКОИДЫ
рин (в коже, а затем — в печени, в стимулированных макрофагах и в почках) при образовании кальциферо ла.
Превращения стероидов состоят в отщеплении алифатической боковой цепи, гидроксилировании и дегидрировании, либо ароматизации А-кольца. Читатель может обратиться к схеме этих процессов, представленной на рис. 3.
Находящийся в составе липосом холестерин и его дериваты претерпевают многократный перенос в митохондрии и эндоплазматический ретикулум, причём наблю-
АНДРОГЕНЫ
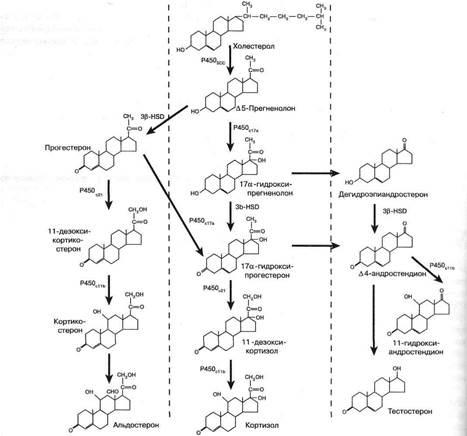
Рис. 3. Пути стероидогснеза в коре надпочечников 14
 дается характерный контакт и «мультипликация» мембранных структур, участвующих в биосинтезе органоидов и липосом.
дается характерный контакт и «мультипликация» мембранных структур, участвующих в биосинтезе органоидов и липосом.
Так как пути стероидогенеза предусматривают участие множества ферментов (например, при продукции эстрогенов — не менее шести), биосинтез стероидов зависит от многих генов, включая как те, которые кодируют ферменты, так и те, которые кодируют внутриклеточные посредники действия стимуляторов стероидогенеза — ключевые белки (Дазо А., 1983). Поэтому существует много наследственных дефектов стероидогенеза, чрезвычайно часто встречающихся в подростковой практике, тогда как число наследственных дефектов продукции пептидных гормонов невелико.
Вместе с тем биосинтез пептидных гормонов нередко происходит эктопически, в опухолевых клетках, так как требует растормаживания всего лишь одной-двух генетических программ. Однако биосинтез стероидов в эктопических опухолях и неэндокринных тканях не типичен.
Стероиды чаще секретируются в готовом виде. Однако возможна и периферическая трансформация прогормонов стероидного происхождения в гормоны. Именно так складывается судьба витамина D в организме. Прогормон, полученный при ультрафиолетовом облучении в клетках кожи или поступивший с пищей, превращается в активную дигидроксифор-му гормона (как уже сказано выше) с участием печени, почек, а иногда — и активных макрофагов.
Почки могут превращать половой гормон прогестерон в минералокортикоид — дезоксикортико-стерон, что имеет значение при развитии отёков у беременных. Плацента переводит дегидроэпиандро- стерон фетальной зоны коры надпочечников плода в эстриол, который позволяет матери избежать вирилизации при беременности мальчиком и участвует в координации сложных процессов формирования эндокринного пола плода. Способность трансформировать некоторые андрогены в эстрогены присуща и трофобластическим опухолям. Липоциты превращают в эстриол андрогены, что имеет большое значение в патогенезе гиноидной формы ожирения (Строев Ю. И. с соавт., 2003). Соответственно как истощение, так и ожирение юношей и девушек могут по-разному сказываться на ходе полового созревания.
Гормоны — производные тирозина
Производные тирозина претерпевают при биосинтезе катехоламинов и дофамина (в мозговом веществе надпочечников, парааортальных ганглиях пло-
да и новорожденного, апудоцитах) гидроксилирование и декарбоксилирование в свободном состоянии и также пакуются в специальные секреторные гранулы.
В щитовидной железе основные этапы превращений тирозина на пути к тироидным гормонам, наоборот, протекают при связывании тирозиновых остатков в составе особого белка — тироглобулина.
Эти предшественники сначала йодируются в двух положениях, образуя монойодтирозин (МИТ) и ди-йодтирозин (ДИТ), затем окисляются и конденсируются между собой, формируя йодтиронины — тетрайодтиронин (тироксин) и трийодтиронин (лиотиронин).
Йодтиронины отщепляются от тироглобулина, который фолликулярные тироциты захватывают из коллоида щитовидной железы, и секретируются в кровь в виде тироксина ( T 4) и трийодтиронина (ТЗ). ТЗ возникает из Т4 путём отщепления йода от наружного фенольного кольца (дейодирование). ТЗ намного более активен, чем Т4, и для него Г, фактически служит прогормоном.
Дейодирование идёт в клетках-мишенях и представляет дополнительный уровень регуляции эффективности тироидной функции, так как может сильно влиять на общую активность тироидных гормонов в разных ситуациях (стресс, голодание, адаптация к переохлаждению и перегреванию и т.п.).
ХРАНЕНИЕ И ВЫДЕЛЕНИЕ ГОРМОНОВ
Хранение и выделение гормонов в биологические среды организма—это важный этап их обмена, объект многочисленных расстройств при патологии. Гормоны выделяются в организме периодически, по потребности в них, и, следовательно, в той или иной мере хранятся в запасе.
Дольше всего хранится неполярный стероид каль циферол. Прекращение поступления и биосинтеза его предшественника у взрослого человека дает эффект в виде остеопороза лишь спустя несколько месяцев, но у ребенка и подростка сказывается значительно быстрее.
Тироглобулин щитовидной железы удерживает двухнедельный запас тироидных гормонов. Его лизис в ходе подострого тироидита Де Кервена может вызывать выбросы в кровь порций гормонов и эпизоды ги-пертироза на фоне общей, свойственной этому заболеванию тенденции к ограничению функциональных возможностей железы — к гипотирозу.
В-клетки островков Лангерганса имеют инсулина не более чем на 5 дней секреции, причём его правильная
15
 упаковка у ряда млекопитающих нарушается блокато-рами цинка, что позволило создать так называемую ди-тизоновую модель сахарного диабета на животных. Другие пептидные гормоны запасаются в еще меньших количествах.
упаковка у ряда млекопитающих нарушается блокато-рами цинка, что позволило создать так называемую ди-тизоновую модель сахарного диабета на животных. Другие пептидные гормоны запасаются в еще меньших количествах.
Кроме кальциферола, другие стероидные гормоны практически не запасаются в силу полярного характера их молекул. Семенники содержат тестостеро на не более чем 15-17 % от суточной потребности, поэтому стероидогенез в них характеризуется высокой постоянной интенсивностью и легко нарушается при острых поражениях яичек, например, при орхите.
Стероидные гормоны освобождаются в кровь постоянно на основе разницы в концентрациях и связываются с липопротеидами и стероидпереносящими белками.
В отличие от них, белковые гормоны и производные тирозина, как правило, поступают в кровь неравномерно. Секреция катехоламинов мозговым веществом надпочечников происходит активно в ответ на симпатический нервный сигнал.
Тироидные гормоны секретируются путем пиноцитоза и протеолиза тироглобулина, которые стимулируются как симпатическими нервами, так и (в основном) гуморальными сигналами — ТТГ и mupocmu - мулирующими иммуноглобулинами. Если печень и другие неэндокринные клетки освобождают свои белки в кровь тонически, мало реагируя на специальные стимулы, то для секреции пептидных гормонов требуется, как правило, активный экзоцитоз секреторных пузырьков. Этот процесс предусматривает пусковой эффект со стороны нервной системы, метаболитов или, чаще всего, других гормонов. Требуется обеспечить работу цитоскелета — нужна затрата макроэргов. И здесь важна роль кальция. Именно так секретируются гормон роста, глюкагон, инсулин. Рис. 4 на примере секреции инсулина дает представление сложности этого процесса.
Секреция инсулина В-клетками островков Лангерганса стимулируется глюкозой, некоторыми аминокислотами (аргинин, лейцин), глюка- гоноподобным полипептидом I и, другими энтериновыми гормонами, особенно желудочным ингиби- рующим полипептидом (ЖИП).
Секреция инсулина стимулируется также холинергическими вагаль-ными и Р-адренергическими симпатическими нервными окончаниями,
16
паракринным глюкагоном, АКТГ и СТГ. В то же время она тормозится соматостатином, а-адренергически-ми влияниями, избытком самого инсулина, амилином, галанином, лептином и многими цитокинами {напри мер, ИЛ-1 и ФНОа). До воздействия секреторных стимулов выход катионов калия поляризует мембрану В-клеток и препятствует входному току ионов кальция через потенциал-зависимые кальциевые каналы. Цитоп-лазматический уровень ионизированного кальция остаётся низким.
При действии секреторного триггера, например, при захвате глюкозы В-клеткой ситуация меняется. Глюкоза метаболизируется, что ведет к генерации АТФ, усилению работы калий-натрий-зависимой АТФ-азы плазматической мембраны, снижению выходного калиевого тока и захвату калия клеткой. Деполяризация приводит к открытию потенциалозависимых входных кальциевых каналов. Катион кальция входит в цитоплазму извне и из внутриклеточных компартментов, где его судьба определяется ц-АМФ-зависимыми механизмами, активированными стимулятором инсулиновой секреции. Кальций влияет на микротрубочки и миозиновые филаменты, способствуя движению цитоскелета и перемещению секреторных гранул с инсулином, который подвергается экзоцитозу с участием белков-кальэлектринов, помогающих слиянию плазматической мембраны и мембран секреторных пузырьков (фъюзогенный эффект).
| Рис. 4. Секреция инсулина В-клетками |
Секреция других пептидных гормонов происходит не менее сложно и, как правило, тесно сопряжена с их биосинтезом, который она активирует. Например, прием глюкозы вызывает быстрый (через 10-15 минут) пик выброса инсулина в кровь, связанный в основном с процессами, описанными выше, но затем этот пик перехо-

 дит в длительное повышение секреции гормона, которое достигает плато примерно через час и вызвано активацией его биосинтеза de novo.
дит в длительное повышение секреции гормона, которое достигает плато примерно через час и вызвано активацией его биосинтеза de novo.
По этим же причинам тройные гормоны активируют как выброс, так и синтез гормонов клеток-мишеней. Например, АКТГ уже через 3-12 минут вызывает выброс в кровь кортикостероидов, а в более отдалённые сроки (1-3 часа) под его влиянием развёртывается картина активации стероидогенеза.
Некоторые гормоны освобождаются в кровь постоянным, меняющимся по интенсивности потоком (па- ратгормон, пролактин, тироидные гормоны). Но для большинства характерен импульсный режим секреции, когда гормон поступает в кровоток дискретными порциями-толчками. Инсулин сочетает и пульсовый, и постоянный режимы освобождения. Но многие другие гормоны — АКТГ, СТГ, гонадотропины, сте роиды — секретируются только импулъсно.
В последнее время обнаружилось, что толчковый дискретный ритм секреции сам по себе несёт сигнальную информацию и важен для действия гормона, а его нарушения свойственны некоторым формам патологии, в том числе — типичным для подростков. Так, люли- берин при пульсовом введении стимулирует продукцию лютеинизирующего гормона (ЛГ) в аденогипофизе, но те же дозы этого гипоталамического рилизинг-фак-тора тормозят продукцию ЛГ, если вводятся в непрерывном режиме. Утрата импульсного характера секреции люлиберина при сохранении его продукции закономерно наблюдается у подростков, страдающих психогенной анорексией-булимией.
Клинический опыт показал, что микроимпульсное введение инсулина (метод «малых доз») более эффективно для контроля уровня глюкозы в крови, чем одномоментная инъекция той же общей дозы. Микроимпульсы поступления СТГ в кровь совпадают с периодами быстрого сна, что наиболее четко выражено у подростков (см. ниже).
В более долговременном плане освобождение гормонов в кровь подчиняется определённым биоритмам (Дедов И.И., Дедов В.И., 1992).
У большинства, хотя и не у всех гормонов, их секреция повышена в ранние утренние часы. Максимальная секреция пролактина осуществляется во время сна. Резкий подъем уровня СТГ у здоровых подростков и молодых людей наступает практически в первый час сна, особенно в период засыпания (дрёмы), что сопровождается знакомым каждому из нас возникающим при засыпании вздрагиванием всего тела (чувство «проваливания»). Зимой многих гормонов (особенно гормонов щитовидной железы) вырабатывается зна-
чительно больше. Тироидных и половых гормонов гораздо больше секретируется в полнолуние, что объясняет романтику лунных ночей.
Под контролем супрахиазматического ядра гипоталамуса, серотонина и мелатонина эпифиза и в зависимости от уровня общей освещённости в видимом диапазоне спектра формируется циркадный (околосу точный) ритм секреции кортиколиберина, а следовательно — АКТГ и глюкокортикоидов. По А. Ша-фарчику с соавт. (1983), у человека и дневных млекопитающих акрофаза секреции гипоталамо-гипо-физарных стимуляторов кортикостероидогенеза приходится на 7-8 часов утра, а минимум — на поздний вечер (20 часов). Установлено, что данный ритм накладывает отпечаток не только на стероидогенез, но и на суточный ход пролиферативных процессов в коре надпочечников (Чурилов Л. П., 1986). Существуют гормональные ритмы с периодом, превышающим сутки {инфрадианные), и более коротким, чем циркад-ные (ультрадианные). Широко известны колебания продукции гонадотропинов и 'половых гормонов, связанные с околомесячным периодом. Они имеют особое значение при установлении месячных у девушек в периоде полового созревания. Не менее важна и околомесячная периодизация эндокринной регуляции у мужчин и юношей.
Инфрадианной периодизации подвержена активность секреции тироидных гормонов. Суммарный результат биоритмологических особенностей продукции гормонов определяет периодические околомесячные и сезонные изменения физической, психоэмоциональной и интеллектуально-поведенческой активности индивидов любого пола, что впервые описано концепциями Г. Свободы-В. Флейса (1898) и А. Л. Чижев ского (1924).
Социально-поведенческая дизритмия является фактором риска многих нейроэндокринных заболеваний. Описаны «болезнь акклиматизации», вызванная быстрой сменой часовых поясов, и учащение артериальной гипертензии у субъектов, занятых ночной работой. В обоих случаях отмечаются аномалии продукции гипоталамических, гипофизарных и надпочечниковых гормонов, гиперфункция ренин-ангиотензин-альдосте-роновой системы.
В подростковом периоде, когда только лишь устанавливаются регуляторные взаимоотношения центрального и периферических звеньев нейроэндокрин-ной системы, такие факторы, как акклиматизация, ночное бодрствование, резкая смена режима дня, перемена часовых поясов — могут быть весьма патогенными. Известен факт, что студенты-подростки чаще страдают гипертонической болезнью при совме-
17
щении учебы с ночной работой. Существующие наблюдения за здоровьем членов юношеских спортивных сборных команд, совершающих международные поездки, это наглядно демонстрируют (Леви на Л. И. с соавт, 1999). При болезни смены часовых поясов отмечено лечебно-профилактическое действие эпифизарного гормона мелатонина и его лекарственных форм — меланекса, мелатонекса. Исчезновение нормального циркадного ритма продукции АКТГ отмечается, например, при болезни Иценко-Кушинга.
Нет сомнений, что биоритмологические подходы, яркое и развёрнутое описание которых можно почерпнуть в учебнике «Патологическая физиология» В. А. Фро лова с соавт. (1999) и в монографии И. И. Дедова и В. И. Дедова «Биоритмы гормонов» (1992), в будущем значительно скажутся на развитии эндокринологии, особенно —ювенильной.
Околосуточный (циркадный) и околомесячный ритмы секреции гормонов должны учитываться при трактовке результатов их исследования. Например, уже сейчас признано, что трактовка данных о содержании тироидных (а половых гормонов — тем более) должна проводиться с учетом того, на какой день месячного цикла брали пробу для анализа у девушек и женщин. Недопустимо, назначив стандартную дозу гормона, игнорировать сезонные и климатические перемены в жизни пациента и связанные с этим колебания потребности в гормонах. Современный эндокринолог, патронируя своих больных на протяжении длительного времени, обязательно должен вносить в дозировку назначенных гормонов коррекции, связанные с биоритмами.
ТРАНСПОРТ ГОРМОНОВ
Транспорт гормонов осуществляется с током крови, лимфы и межклеточной жидкости. После секреции гормоны попадают в равновесную систему, где концентрации связанного и свободного биорегулятора в крови соотносятся между собой и зависят от баланса скорости поступления и скорости удаления гормона из крови, а также от количества и аффинности белка-переносчика. Гормоны быстро разрушаются, однако, постоянно синтезируясь, находятся у здоровых в количествах, потребных для сохранения гоме-остаза.
Деградация гормонов происходит в тканях-эффекторах, а также в печени и почках, продукты их распада выделяются в основном с мочой и желчью. О скорости использования гормонов судят по их клиренсу (Т1/2). В клинике о продукции гормонов можно сделать заключения по их концентрации в биологических жидкостях, оп-
18
ределяемой иммуноферментными, иммунохемилюми-несцентными или радиоиммунологическими (реже — биохимическими или биологическими) методами. Однако надо помнить, что концентрации (содержание, уровень) гормонов в крови — это параметры динамические, равновесные. В связи с этим очевиден архаизм традиционно неточных терминов—«гормональный статус (фон, зеркало)», которые авторы настоящего руководства никак не могут рекомендовать к употреблению.
У пептидных гормонов время их полужизни в крови — порядка 3-7 минут, а у гликопротеидных оно может быть более часа, но остается для всех этих гормонов принципиально весьма коротким, что позволяет эндокринной системе оперативно менять гормональный уровень.
Пептидные гормоны распространяются в свободном виде, но тироидные и стероидные гормоны в силу их гидрофобности требуют солюбилизаторов-перенос-чиков. Наиболее известные транспортёры гормонов — транстиретин (преалъбумин), переносящий тироид ные гормоны и ретинол, а также такие белки, как: ти-роксин-связывающий глобулин, тестостерон-связы- вающий глобулин, кортикостероид-связывающий глобулин, альбумин, который неспецифически фиксирует тироидные и многие стероидные гормоны. Сте роиды и тироидные гормоны могут также переноситься липопротеидами, особенно липопротеидами высокой плотности. Тироидные гормоны за счет своих антиок-сидантных свойств препятствуют атерогенным окислительным изменениям липопротеидов. Под их влиянием снижается экспрессия высокоатерогенных липопротеидов (Дантес Л. Г., 2002).
Многие эндокринологические руководства к важным причинам эндокринопатий традиционно причисляют нарушения транспорта гормонов (по аналогии с фармакологическими данными о том, что связывание с белками крови очень существенно для определения биодоступности лекарств). Однако Дж. Уилсон (1994) справедливо указывает, что при интактной обратной связи в системе «гипоталамус — гипофиз — периферические железы» сдвиги концентраций свободного активного гормона, наступающие от изменения связывания с плазменными белками, не будут иметь стабильного характера. Ведь у здорового индивида они эффективно нейтрализуются компенсаторным изменением секреции гормона.
Так, эстрогены увеличивают содержание тирок- син-связывающего глобулина и посему делают биодоступность тироксина меньшей. В ответ на это у женщин в эстральную фазу месячного цикла гипота-
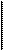
 ламо-гипофизарный нейросекреторный аппарат по принципу обратной связи увеличивает продукцию ти- ролиберина и ТТГ, а щитовидная железа добавляет в кровь тироидные гормоны — и эффективная биодоступная концентрация этих регуляторов остается нормальной. В плазме наступает кажущееся увеличение концентрации общего Т4, не означающее наступления функционально-метаболического гипертироза. Однако у женщин это небезразлично при истолковании данных о концентрациях тироидных гормонов в их крови. То же имеет место при беременности и при пользовании гормональными контрацептивами.
ламо-гипофизарный нейросекреторный аппарат по принципу обратной связи увеличивает продукцию ти- ролиберина и ТТГ, а щитовидная железа добавляет в кровь тироидные гормоны — и эффективная биодоступная концентрация этих регуляторов остается нормальной. В плазме наступает кажущееся увеличение концентрации общего Т4, не означающее наступления функционально-метаболического гипертироза. Однако у женщин это небезразлично при истолковании данных о концентрациях тироидных гормонов в их крови. То же имеет место при беременности и при пользовании гормональными контрацептивами.
Иное дело, когда имеются отклонения от нормы во взаимоотношениях центрального и периферического звеньев нейроэндокринной системы. При нарушении нормальных механизмов сервоконтроля гормональных концентраций колебания степени связывания гормонов в крови становятся существенным патогенетическим фактором эндокринопатий. Так, даже небольшие дозы глюкокортикоидов, вводимых с лечебной целью, способны вызывать синдром Кушинга у подростков с ги-попротеинемическими состояниями и со снижением продукции кортикостероид-связывающего белка (при недоедании, печеночной недостаточности, нефротичес-ким синдроме, микседеме).
ИНАКТИВАЦИЯ ГОРМОНОВ
Инактивация гормонов — важный этап их метаболизма. Каждый, кому случалось иметь дело с запавшей кнопкой дверного звонка, помнит, как надоедливо звучит его непрекращающийся сигнал. Химический сигнал-гормон, как и любой другой, для эффективного использования должен своевременно прекращать своё действие — инактивироваться.
Выше уже упоминалось, что гормоны инактивиру-ются как в клетках-мишенях после их проникновения во внутриклеточное пространство, так и в нетаргетных органах, главным образом, в печени и в почках. Иммунные комплексы с участием гормонов подвергаются клиренсу в фагоцитах.
Основной путь инактивации пептидных гормонов — протеолиз. Хотя исторически сложились представления о существовании для некоторых гормонов специальных протеаз, на деле эти процессы осуществляются в основном неспецифическими протеолитичес-кими ферментами клеток-мишеней. Так, до сих пор еще не обнаружено специальной «инсулиназы», хотя условно этим понятием оперируют многие эндокринологические руководства. Вместе с тем для многих па-ракринных пептидов — например, интерлейкинов,
кининов, ангиотензинов — имеются специальные кон-вертазы (например, интерлейкин-конвертаза ICE, анги-отензин-конвертаза АСЕ, калликреин).
Гормоны тирозинового происхождения инакти-вируются специальными ферментами. Катехолами-ны разрушаются моноаминоксидазой (МАО) и кате-хол-ортометилтрансферазой (КОМТ) тканей-мишеней и печени. Окислительное дезаминирование и орто-ме-тилирование этих гормонов ведет к образованию и к экскреции с мочой таких продуктов, как ванилил-мин-далъная кислота, норметанефрин и метанефрин, концентрации которых определяют в диагностических целях в моче. Блокада ферментов катаболизма кате- холаминов, например, с помощью лекарств приводит к значительному увеличению чувствительности тканей к катехоламиновым сигналам.
Тироидные гормоны дейодируются в тканях-мишенях и печени, дезаминируются и деконденсируют-ся в печени, а затем окисляются, выводятся в кишечник с желчью и оттуда в небольших количествах — рециркулируют.
Стероиды подвергаются в печени восстановлению (для чего имеются специальные энзимы), затем — неспецифическому гидроксилированию. Наконец, они образуют гидрофильные парные соединения с активными формами серной или глюкуроновой кислоты и поступают в желчь и мочу. Из желудочно-кишечного тракта их дериваты могут реабсорбировать-ся, а содержание продуктов распада стероидов в моче (например, 17-КС, 17-ОКС) измеряют в диагностических целях.
Как указывалось выше, в связи с закономерностями гормонального транспорта сами по себе колебания скорости деградации гормонов напрямую не приводят к эндокринной патологии, если только функционирует сервостатический механизм гипота-ламо-гипофизарного контроля над их эффективными концентрациями. Может быть, единственным примером такого рода патологии служит резистентность к экзогенному инсулину из-за его аномально быстрого расщепления при подкожных инъекциях.
Вместе с тем надо помнить, что при патологии обратные связи в системе могут быть нарушены, и клиренс гормонов становится важной детерминантой их эффективной концентрации. Так, гипертироз ускоряет катаболизм стероидов, делая подростков с базедовой болезнью относительно «резистентными» к глю-кокортикоидам. Печеночная недостаточность замедляет клиренс алъдостерона, серотонина, эстрогенов, кортизола и других гормонов, что на-
19
 кладывает отпечаток на симптомы, наблюдаемые у больных. Скажем, такие клинические проявления циррозов печени у мужчин, как импотенция, гинекомастия, отёки, эритема ладоней и сосудистые «звёздочки» объясняют избытком соответствующих гормонов, недоразрушенных плохо функционирующими гепато-цитами.
кладывает отпечаток на симптомы, наблюдаемые у больных. Скажем, такие клинические проявления циррозов печени у мужчин, как импотенция, гинекомастия, отёки, эритема ладоней и сосудистые «звёздочки» объясняют избытком соответствующих гормонов, недоразрушенных плохо функционирующими гепато-цитами.
МЕХАНИЗМЫ ДЕЙСТВИЯ ГОРМОНОВ
И ВАЖНЕЙШИЕ ПРИНЦИПЫ ЭНДОКРИНОЛОГИИ :
Дата: 2019-02-02, просмотров: 345.