Кишечник. На рисунке показана типичная пейерова бляшка подвздошной кишки. Подобные лимфоидные образования расположены по всему кишечнику и реагируют на антигены, поступающие через кишечную стенку. Их называют лимфоидной тканью, ассоциированной с кишечником. В ней содержатся специализированные М-клетки, транспортирующие антиген из просвета кишечника в субэпителиальный слой, где присутствуют в большом количестве лимфоциты, особенно CD8+ Т-клетки с гамма/дельта-pe-цептором. Далее в собственной пластинке представлены все типы Т- и В-лимфоцитов, особенно IgA В-лимфоциты, макрофаги и тучные клетки. Только в 2 м кишечника содержится столько же лимфоцитов, сколько во всём костном мозге, лимфатических узлах и селезёнке вместе взятых.
Лёгкие. Так же как кишечник, лёгкие являются главным органом контакта с внешними антигенами и имеют сходную организацию лимфоидной ткани в ассоциации с бронхами.
Глотка. Лимфоидные образования (миндалины, аденоиды) реагируют на антигены, поступающие через носоглотку. Имеют зародышевые центры и Т- и В-лимфоциты. Слюнные железы содержат также лимфоциты, происходящие из ЛТАС.
Кожа. Антигены, проникшие через кожу, захватываются клетками Лангерганса (см. также рис. 7) и транспортируются в Т-зону лимфатического узла. Эти клетки чрезвычайно чувствительны к ультрафиолетовому излучению, которое снижает реакции контактной гиперчувствительности и способствует индукции, в большей степени супрессорной, чем хелперной, функции Т-клеток (значение этого феномена при раке кожи представлено на рис.31).
29
12. ЭВОЛЮЦИЯ РАСПОЗНАЮЩИХ МОЛЕКУЛ
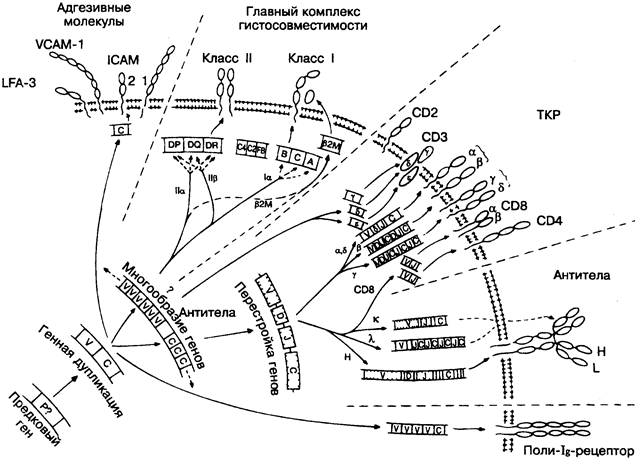
В начале этой главы уместно напомнить о различии между естественным и адаптивным иммунитетом, которое наиболее существенно в разрешающей способности распознающих их структур.
Естественное иммунное распознавание, например фагоцитами, клетками-киллерами или по альтернативному пути активации комплемента, которое ещё недостаточно изучено, по-видимому, основывается на относительно простом механизме различения. Обычно частный чужеродный материал распознаётся и удаляется или не распознаётся (эффект по принципу «друг или враг»).
Распознавание лимфоцитами (основными клетками адаптивного иммунитета) происходит по-другому. Огромный спектр чужеродных субстанций различается индивидуально, после чего запускается соответствующий вариант ответа на них. Этот процесс стал возможным в результате эволюции 3 серий клеточно-поверхностных рецепторов с высочайшей степенью гетерогенности, а именно молекулы антитела, Т-клеточного рецептора и молекулы главного комплекса гистосовместимости. Благодаря работам в области молекулярной биологии выяснилось, что все эти рецепторы имеют столько сходных последовательностей на уровне гена (ДНК) и белка (аминокислоты), что мож-
но уверенно говорить о их происхождении из общего предшественника — вероятно, одной из распознающих молекул ранних ступеней эволюции (рис. 3). Так как антитела были первыми идентифицированы в этой генетической системе, то они в совокупности часто обозначаются как суперсемейство иммуно-глобулиновых генов, куда вошли и другие близкие по свойствам молекулы. Некоторые из них действительно выполняют иммунные функции. Характерная особенность всех этих молекул — домены, представляющие собой бетта-складчатые структуры из свёрнутых последовательностей примерно 110 аминокислот (изображены на рисунке в виде округлых петель, выступающих из клеточной мембраны).
Предстоит большая исследовательская работа, чтобы заполнить белые пятна в эволюции представленных молекул. Рисунок лишь иллюстрирует вероятную взаимосвязь между молекулами этого удивительного семейства. Современные знания о их функциях и строении изложены в следующих 4 рисунках.
Гипотетический ген-предшественник суперсемейства иммуноглобулиновых генов не обнаружен ни у одного из существующих биологических видов, но, возможно, кодировал первые распознающие молекулы, например у губок (рис. 3).
30
Гены V, С. Первым жизненно важным шагом была дупликация предковсго гена (P-гена) в два гена, один из которых дал начало всем существующим современным вариабельным генам, а другой — константным генам. На рисунке представлены гены и полипептиды с достаточной гомологией, чтобы считаться частью семейства вариабельных генов. Последующие дупликации со всё большей изменчивостью вариабельных и константных генов привели в итоге к большому разнообразию наблюдаемых сегодня доменов.
Главный комплекс гистосовместимости (ГКГС) представлен на рисунке как система HLA генов (лейкоцитарные антигены человека). Она кодирует два типа молекул. Гены ГКГС класса I кодируют поверхностные молекулы, присущие всем ядросодержащим клеткам, а гены ГКГС класса II — поверхностные молекулы, присущие только некоторым иммунным клеткам. Известно. что альфа- и бетта-цепи молекул ГКГС класса II содержат константные и, возможно, вариабельные регионы. Взаимодействие молекул ГКГС и Т-клеточных рецепторов необходимо для всех адаптивных иммунных реакций. Число генных локусов ГКГС очень различается у разных биологических видов. У человека, например, их 6, что довольно близко к оптимальному значению. Интересно, что в процессе метаморфозы амфибий сначала появляются молекулы класса II, а затем — класса I.
Бетта2-Микроглобулин (бетта2М), присоединяясь к цепям класса I, завершает образование четырёхдоменной молекулы. Предшественник бетта2М ещё не выявлен.
Три компонента комплемента — С2, С4, FB — кодируются генами в ГКГС, но структурно не связаны с его молекулами.
Перестройка генов — процесс, характерный только для Т- и В-лимфоцитов. Последовательные вырезания участков ДНК и информационной РНК приводят к тому, что молекула каждого рецептора кодируется сочетанием V-, J- (и О- для цепей IgH)-генов, каждый из которых выбран из набора, содержащего от 2 до 100 и более генов, что в сумме даёт тысячи возможных комбинаций. Так как уникальная перестройка гена наблюдается в каждой Т- и В-клетке и затем наследуется их потомками, каждый лимфоцит или клоны лимфоцитов строго индивидуальны, что создаёт основу для всех адаптивных иммунных реакций (рис. 17-20).
Т-клеточный рецептор (ТКР) — комплекс поверхности молекул Т-клеток, включающий альфа- и бетта- или гамма- и дельта-цепи ТКР, CD3,
CD4 или CD8 молекулы в зависимости от типа Т-клетки. Все они образуют комплекс, при помощи которого Т-лимфоцит распознаёт специфический антиген вместе с молекулой ГКГС, чтобы активироваться и выполнять свои функции (хелперные, цито-токсические и др.).
Антитела. Антитела или иммуноглобулиновая молекула составляют часть клеточно-поверхностного рецептора В-лимфоцитов или секретируются в огромном количестве активированными В-лимфоцитами и создают комплекс сывороточных антител, защищающих в основном от патогенных микроорганизмов. Домены практически сходны с доменами альфа- и бетта-цепей ТКР, но собраны по другому принципу: две четырёхдоменные тяжёлые (Н) цепи связаны с двумя двухдоменными лёгкими (L) цепями.
Следует отметить, что формирование разнообразия в генах для различных цепей не всегда происходит по данному сценарию. Например, тяжёлые и лёгкие (к) цепи млекопитающих вместе имеют J-гены между V- и С-генами, а лёгкие лямбда-цепи имеют повторные J-C-сегменты (см. рисунок). У акул дуплицируется сразу весь сегмент VDJC, что обеспечивает гораздо меньшую вариабельность.
Поли-Ig-рецептор — молекула, обнаруженная на некоторых эпителиальных клетках, способствующих транспорту антител в секреты, например, в слизь. Многие другие молекулы, включая некоторые Fc-рецепторы, адгезивные молекулы (см. ниже), рецепторы к факторам,роста и цитокинам, имеют частичную характерную доменную структуру. Их общая характеристика — вовлечение в межклеточные взаимодействия. Для антител это скорее исключение, чем правило.
Адгезивные молекулы — поверхностные молекулы, о которых упоминалось в разделе о воспалении (рис. 6). Усиливая соединение клеток, они облегчают межклеточную кооперацию и прикрепление к стенкам кровеносных сосудов. Некоторые адгезивные молекулы принадлежат к суперсемейству иммуно-глобулинов и обычно связаны с одним или несколькими соответствующими лигандами, например:
CD2..............................LFA-3
VCAM-1.......................VLA-4
ICAM-1, ICAM-2, ICAM-3 .....................LFA-1
31
13. ГЛАВНЫЙ КОМПЛЕКС ГИСТОСОВМЕСТИМОСТИ (ГКГС)
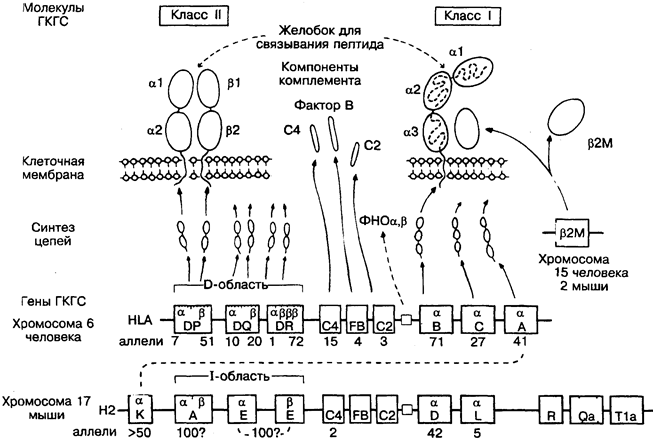
Большая и важная группа генов, получивших в своё время не совсем благозвучное определение на основании того, что белки, кодируемые ими, были впервые выявлены при отторжении трансплантата, т.е. при тканевой несовместимости. Ныне установлено, что истинная роль этих белков — служить маркёрами идентичности на поверхности различных клеток, с которыми Т-лимфоциты взаимодействуют через собственные рецепторы в иммунной реакции.
Исторически сложилось, что у мышей ГКГС известен как Н2, а у человека — как HLA ( human leucocyte antigens}. Изучение ГКГС у разных биологических видов пока не выявило существенных отличий по структуре и механизмам взаимодействия с Т-клетками. ГКГС состоит из группы генов класса 1 и класса II. На рисунке обозначения генов заключены в прямоу-
гольники, под которыми указано число аллелей (альтернативных вариантов) для каждого локуса. ГКГС обладает огромным полиморфизмом, и только в пределах 1 хромосомы число возможных комбинаций её генов превышает 3-106. Молекулы ГКГС, кодируемые обеими хромосомами, создаются на основе 1 из почти 1013 комбинаций генов, что неимоверно осложняет пересадки органов, например почки (рис. 37).
Когда типирование HLA-системы получило широкое распространение, выяснилось, что от её состава зависит предрасположенность или, напротив, повышенная резистентность к многим заболеваниям. Было предложено несколько объяснений этой связи, но ни одно из них не является достаточно убедительным.
32
Н2 — мышиный ГКГС, расположенный на хромосоме 17. По крайней мере 20 других генов минорной гистосовместимости расположены на разных хромосомах и обозначены как HI, H3 и т.д., но только Н2-гены оказывают очень существенное влияние на отторжение трансплантата. Установлено также, что лишь они кодирует молекулы для нормальных межклеточных взаимодействий.
К, D, L — гены Н2 класса I, кодирующие альфа-цепь (м. м. 44 000), которая в сочетании с бетта2-микроглобулином образует четырёхдоменные молекулы К, D или L или антигены. N-концевые участки альфа-цепей крайне вариабельны и, возможно, эволюционировали для взаимодействия с различными вирусами. Поскольку вирусом может быть заражена любая клетка, то молекулы ГКГС класса I присутствуют на поверхности практически всех клеток организма, за исключением эритроцитов у некоторых биологических видов (в том числе у человека). На рисунке под каждым локусом проставлено число известных аллелей. Фактически их гораздо больше.
R, Qa, Tla считаются частью семейства класса I, так как кодируют очень сходные молекулы, которые представлены только на некоторых лимфоцитах и не опосредуют отторжение трансплантата.
А, Е — гены Н2 класса II, объединённые в l-область. А и Е содержат разные гены для альфа- (м. м. 33 000) и бетта- (м. м. 28000) цепей четырёхдоменной молекулы. В отличие от молекул класса I, молекулы класса II присутствуют только на клетках, взаимодействующих с Т-лимфоцитами и регулирующих их. Вероятно, огромное разнообразие молекул класса II объясняется существованием большего, чем показано на рисунке, количества генов А и Е со многими аллелями.
HLA — ГКГС человека, расположенный на хромосоме 6. Аналогичен Н2, за исключением того, что гены класса I расположены вместе и существуют как минимум 3 гена класса II.
А, В, С — гены HLA класса I, из них А — гомолог гена К у мышгй. Существуют также гены HLA Е, F, G и Н, но функции их продуктов пока неясны.
DP, DQ, DR — гены HLA класса II. Продукты DP гена весьма эффективно стимулируют пролиферацию Т-клеток и выявляются в смешанной реакции лимфоцитов, тогда как все другие антигены классов I и II определяются антисыворотками. В этой же области находятся и другие гены (DN, DO, DZ), продукты которых пока неизвестны. Почти всю вариабельность,
необходимую для взаимодействия с различными антигенами, а затем и с Т-клеточным рецептором, в молекулу класса II вносит бетта-цепь.
Обычно активно захваченный клеткой и расщеплённый в лизосомах до пептидов антиген связывается с молекулой ГКГС класса II, а антиген, образованный внутри клетки (например, из вируса), связывается с молекулой ГКГС класса I. Более подробно о взаимодействии антигенных фрагментов и молекул ГКГС см. на рис. 14 и 17.
С2, С4, FB — гены ГКГС класса III. Кодируют компоненты комплемента, участвующие в активации СЗ. Интересно, что, хотя все они существуют в нескольких аллельных формах, значимых последствий этого не обнаружено. В этой же области находятся гены, кодирующие фермент надпочечников 21-гидроксилазу, цитокины ФНО альфа и бетта. Молекула, кодируемая С4, способна присоединяться к эритроцитам и мимикрировать под антигены группы крови.
бетта2М — бетта2-микроглобулин (м. м. 12 000). Кодируется не в ГКГС, но входит в состав всех молекул класса I, стабилизируя их на поверхности клеток. В сыворотке обнаруживается в растворимой форме. У мышей имеет 2 аллельные формы, но в общем это одна из наиболее известных консервативных молекул.
Дата: 2018-12-28, просмотров: 460.