1) Периферический отдел. Боль возникает в результате воздействия раздражителей на спец. высокопороговые болевые рецепторы, реагирующие на стимулы, угрожающие организму повреждением или вызывающие повреждения. Основные ноцицепторы — механоцепторы тонких миелинизированных волокон (Аσ-механорецепторы) и полисенсорные ноцицепторы немиелинизированных С-волокон (С-ноцицепторы реагируют на механические, химические и температурные раздражители). Аσ-ноцицепторы обычно не реагируют на термические, химические и болевые раздражители, но возбуждаются при сильных механических воздействиях (щипок пинцетом, укол иглой).
Болевые рецепторы всех органов и тканей представляют неинкапсули-рованные окончания нервных волокон, имеют форму волосков, пластинок, сплетений, спиралей.
Основная часть ноцицепторов кожных, мышечных и внутренних органов полисенсорные. Моносенсорные ноцицепторы (механо- и термоцепторы) встречаются редко. «Быстрая» боль вызывается механ. и t-ными стимулами. «Медленная» боль вызывается всеми видами стимулов.
Полагают, что при действии механического раздражителя РП возникает в результате деформации и растяжения мембраны, что активирует Na-каналы и происходит диффузия Na+ внутрь ноцицептора. Возможно, что также под действием механического раздражителя из терминалей сенсорной клетки выделяется медиатор (субстанция П). В последующем он реагирует со специфическими хеморецепторами мембраны ноцицептора, запуская генерацию РП.
В случае термического воздействия, формирование РП протекает, очевидно, также с участием медиатора, который затем вступает в реакцию с хеморецепторами этого же рецептора, запуская генерацию РП, при этом крутизна и амплитудные характеристики определяются параметрами t-ного воздействия. Медиаторами болевых первичных нейронов является глутамат (модуляторами — субстанция Р, ВИП).
При воспалительных процессах ощущение боли возникает под действием тканевых алгогенов — веществ, появляющихся в результате разрушения тканевых структур (гистамин, серотонин, ионы К+, эйкозаноиды — простагландины и лейкотриены), а также алгогенов плазмы крови, появляющихся в результате выхода в межклеточное пространство компонентов крови, катализирующих механизм образования кининов (каллидина и брадикинина). Они открывают ионные каналы либо активируют системы вторых посредников и деполяризуют (возбуждают) неповрежденные соседние рецепторы. При этом галогены действуют и на ноцицептор, и на нервное окончание.
 При действии болевого раздражителя или воспаления обычно наблюдается сенситизация ноцицепторов — повышение их возбудимости, вследствие чего резко возрастает болевая чувствительность на повторное раздражение. Алгогены всех типов отчетливо угнетают клеточное дыхание, что также способствует формированию болевых ощущений.
При действии болевого раздражителя или воспаления обычно наблюдается сенситизация ноцицепторов — повышение их возбудимости, вследствие чего резко возрастает болевая чувствительность на повторное раздражение. Алгогены всех типов отчетливо угнетают клеточное дыхание, что также способствует формированию болевых ощущений.
2) Проводниковый отдел начинается в основном тонкими немиел. волокнами типа С (v проведения возбуждения = 0,5-2 м/с, d=0,5-1 мкм), а также миел. волокнами типа Аσ (v=5-15 м/с, d=1-4 мкм).
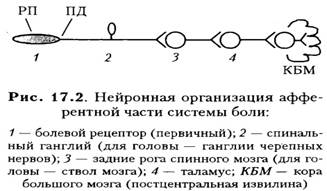
Болевая импульсация от тела организма поступает по Аσ-и С-волокнам — это дендриты биполярных нейронов спинальных ганглиев (1ые нейроны), они вступают в сп. мозг в составе задних корешков. Аксоны этих нейронов в сп. мозге переключаются в задних рогах (2ые нейроны) и переходят на противоположную сторону.
- Окончания С-волокон, контрактирующие с нейронами сп. мозга, выделяют 2 медиатора – глутаминовую кислоту и субстанцию П. Глут. кислотадействует быстро, реализуя свой специф. эффект в течение нескольких мс, тогда как эффект субстанции П реализуется медленно из-за того, что наращивание её концентрации осущ. в течение несольких секунд или даже минут.
С-волокна направляются в ствол мозга в составе латерального спиноталамического тракта и на уровне ствола мозга образуют синапсы с нейронами серого околоводопроводного вещества ядра тектальной области, ядра РФ продолг. мозга, моста, среднего мозга. Многократно переключаясь на нейронах РФ, возбуждение в последующем направляется преимущественно в неспециф. ядра таламуса, частично — в специфические, а также направляется в структуры гипоталамуса и подкорковые структуры.
- Аσ-волокна направляются в ствол мозга в составе переднего спиноталамического тракта, вступают в таламус, где происходит переключение на нейроны специфического сенсорного комплекса ядер вместе с волокнами дорсальных столбов (3-и нейроны), формирующих медиальную петлю с другими волокнами. На своем пути в стволе мозга А8-волокна отдают коллатерали к ядрам стволовой РФ.
Болевая импульсация от кожи лица и органов челюстно-лицевой области поступает по Аσ- и С-волокнам, которые в составе тройничного нерва (аксоны первых нейронов — в ганглии тройничного нерва, расположенного на височной кости) направляются в продолг. мозг, где заканчиваются на нейронах ядра спинального тригеминального тракта (2ые нейроны). Часть волокон подходит к ядрам РФ продолг. мозга, а также к ядрам шва, далее — к нейронам специф. и, частично, неспециф. ядер таламуса (3и нейроны).
3) Корковый отдел локализуется, в основном, в первичной соматосенсорной проекционной коре(поле S1), обеспечивающие восприятие «быстрой» боли. Точная идентификация участка кожной поверхности, на которую действует болевой раздражитель, требует параллельного вовлечения в процесс кожных тактильных рецепторов. Полю S, отводится ведущая роль в экстренном включении моторной защитной реакции организма в ответ на действие болевого раздражителя. От поля S, импульсация поступает к нейронам моторной коры по прямым синаптическим связям. Ощущения «медленной» боли обычно идентифицируются плохо, пути ее проведения представляют в основном полисинапт. нейронные цепи РФ, неспециф. ядер таламуса, генерализованно передающих возбуждение в различные корковые поля, в том числе в поля S1 и S2 болевой системы. Вторичное соматосенсорное проекционное поле S2 располагается на границе пересечения центр. борозды с верхним краем височной доли. Нейроны поля S2 имеют двусторонние связи с ядрами таламуса, что позволяет данному полю избирательно фильтровать проходящее через таламус возбуждение, и прежде всего болевого происхождения. После такого выделения поле S2 вовлекается в процессы деятельности мозга, связанные с извлечением из памяти энграммы нужного поведенческого акта, и в формирование ответной реакции организма.
Дата: 2019-07-30, просмотров: 1885.