Механизмы оценки характера звука. Различение высоты звуков. Характер и локализация колебаний базальной мембраны меняется в зависимости от частоты звука. Так, звуки низкой частоты приводят в колебательные движения всю мембрану, но особенно сильно в области, ближайшей к геликотреме — верхнему отделу улитки, где находятся более длинные волокна основной мембраны.
При действии тонов до 400 Гц осуществляется пространственно-частотное кодирование.Пространственное кодирование заключается в том, что информация в виде нервных импульсов передается по определенным волокнам слухового нерва. Частотное кодирование выражается тем, что частота афферентных импульсов в слуховом нерве повторяет частоту звуковых колебаний, поскольку она не превышает лабильность этих нервных волокон.
С увеличением частоты колебаний звуковых волн выше 400 Гц максимальная амплитуда колебаний основной мембраны смещается к нижней части улитки (ближе к овальному окну), где расположены более короткие волокна. При этом частота возникающих в слуховом нерве ПД не возрастает. Характер импульсаций меняется в соответствии с местом наиболее интенсивных колебаний базальной мембраны (теория места). Первичная слуховая кора также топографически поделена на области, которые отвечают за восприятие звуков различной высоты: низкочастотные звуки воспринимаются передними отделами височной коры. Нейроны задних отделов слуховой коры воспринимают информацию о высокочастотных колебаниях. Таким способом осуществляется пространственное кодирование высоты звуковых сигналов, частота которых выше 400 Гц, что согласуется с резонансной теорией Гельмгольца. Др. теории:
- Телефонная теория Резерфорда: частотно кодирование (до 1000 Г).
- Комбинированная теория – теория бегущей волны.
Известны три основных механизма различения силы звуковых колебаний:
1) с увеличением силы звука увеличивается амплитуда колебаний базальной мембраны, что ведет к возрастанию амплитуды РП и, следовательно, частоты ПД, распространяющихся по слуховому нерву в корковый отдел системы слуха;
2) чем звук сильнее, тем больше площадь колеблющейся базальной мембраны и соответственно большее число волосковых клеток одновременно вовлекается в колебательный процесс; увеличение рецептивного поля приводит к пространственной суммации в ЦНС;
3) в возбуждение вовлекаются менее возбудимые и более глубоко расположенные в кортиевом органе рецепторы (внутренние клетки). Чем громче звук, тем вовлекается больше наружных клеток.
Определение локализации источника звука возможно с помощью бинаурального слуха. Бинауральный слух (способность слышать одновременно двумя ушами) обеспечивает сравнение характеристик звуковых колебаний, полученных каждым ухом. Звуковая волна раньше поступает к уху, на стороне которого локализуется источник звука: оценивается разность во времени прихода одинаковых фаз звуковой волны к обоим ушам. Т. о., бинауральный слух, по сравнению с моноауральным, обеспечивает более точное определение локализации источника и соответственно направления звука.
11. Особенности организации вестибулярного анализатора, его роль в поддержании равновесия, положения тела в пространстве.
Вестибулярная система обеспечивает восприятие информации о прямолинейных и вращательных ускорениях движения тела и изменениях положения головы в пространстве или всего тела, а также о действии земного тяготения. У этой системы практически нет адаптации.
1) Периферический (рецепторный) отдел представлен двумя типами рецепторных волосковых клеток вестибул. органа, расположенного вместе с улиткой в лабиринте височной кости. Вестибул. орган состоит из преддверия (сферич. и эллптический мешочек) и трех полукружных каналов, которые открываются своими устьями в преддверие. Один из концов каждого канала расширен в виде ампулы. Все эти структуры образуют перепончатый лабиринт, заполненный эндолимфой. Между перепончатым и костным лабиринтом находится перилимфа. В мешочках преддверия имеются небольшие возвышения - пятнами (макулами), а в ампулах полукружных каналов — гребешками (кристами). Здесь находятся рецепторные эпителиальные клетки, имеющие на свободной поверхности тонкие, многочисленные (40 — 60 шт.) волоски (стереоцилии) и один более толстый и длинный волосок (киноцилию).
Рецепторные клетки преддверия покрыты отолитовой мембраной (желеобразной массой из мукополисахаридов с кристалликами карбоната кальция — отолитов, а находящиеся в ампулах — листовидной мембраной (без отолитов). В эти мембраны погружены волоски рецепторных клеток. Возбуждение волосковых клеток происходит при изгибании стереоцилий в сторону киноцилий и открытия механочувствит. калиевых каналов. Калий согласно градиенту концентрации поступает из эндолимфы в цитоплазму и деполяризует мембрану. Возникающий РП вызывает выделение из волосковых клеток АХ, который действует на холинорецепторы постсинаптической мембраны окончания дендрита нервного волокна первого нейрона вестибулярной системы. На постсинаптической мембране возникает ГП, эл. поле которого обеспечивает возникновение ПД в афферентном волокне вестибулярного нерва. При смещении стереоцилий в противоположную от киноцилий сторону происходит закрытие ионных каналов, гиперполяризация мембраны и понижение активности сенсорных волокон вестибулярного нерва.
Адекватным раздражителем для рецепторных клеток преддверия являются линейные ускорения и наклоны головы или всего тела, приводящие к скольжению отолитовых мембран и изгибанию волосков. Для рецепторных клеток ампул полукружных каналов адекватным стимулом являются угловые ускорения в разных плоскостях при поворотах головы или вращении тела. В начале угловых ускорений деформация ресничек обусловлена смещением рецепторных клеток относительно временно неподвижных листовидных мембран из-за инерции эндолимфы, а при завершении движения — скольжением листовидной мембраны при уже неподвижном рецепторе.
2) Проводниковый отдел представлен, афф. (сенсорными), так и эфф. (меняющими функц. мобильность волосковых клеток) волокнами. Под влиянием медиатора на постсинапт. мембране дендрита биполярного нейрона возникает ГП, обеспечивающий действием своего эл. поля возникновение ПД биполярного нейрона. Аксоны биполярных нейронов (1ый нейрон вестибулярного пути) вестиб. ганглиев направляются к вестибулярным, ядрам (Бехтерева, Роллера, Швальбе и Дейтерса) продолговатого мозга. Нейроны (2ой нейрон) этих ядер получают доп. информацию от рецепторов мышц и суставов (шеи и др.), 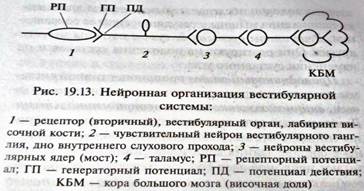 сетчатки и мышц глаза, мозжечка и сами посылают сигналы ко многим другим отделам ЦНС: спинному мозгу; глазодвигательным ядрам; вестибулярным ядрам противоположной стороны мозга; мозжечку; ретикулярной формации; гипоталамусу; таламусу, в постлатераль- ном ядре которого расположены 3и нейроны, аксоны которых образуют прямой путь в кору большого мозга.
сетчатки и мышц глаза, мозжечка и сами посылают сигналы ко многим другим отделам ЦНС: спинному мозгу; глазодвигательным ядрам; вестибулярным ядрам противоположной стороны мозга; мозжечку; ретикулярной формации; гипоталамусу; таламусу, в постлатераль- ном ядре которого расположены 3и нейроны, аксоны которых образуют прямой путь в кору большого мозга.
3) Корковый отдел вестиб. системы (первая вестиб. зона) представлен нейронами (4ые нейроны), рассеянными в коре теменной и височной долей — в постцентр. извилине, верхней височной извилине, верхней теменной дольке, поля 20 и 21. В премоторной коре обнаружена вторая вестибулярная зона, в которую поступает информация по непрямому вестибулоцеребеллоталамическому пути.
3 типа вестибулярных реакций:
Соматические реакции обеспечивают поддержание тонуса мышц, координацию движений, нужное положение тела в пространстве и соответствующие глазодвигательные реакции. К ним относятся многие врожденные (безусловные) спинальные (через вестибуло- и ретикулоспинальные тракты к у- и/или а-мо- тонейронам спинного мозга) и стволовые (связи вестибулярных ядер с мотонейронами глазодвиг. ядер) рефлексы — статические, статокинетические и кинетические. При этом вертикальная поза и походка определяются преимущественно отолитовым аппаратом, а глазодвигательные реакции вызываются с рецепторов полукружных каналов. Вестибулярный нистагм — особая форма глазодвигательных реакций, необходимая для удержания объекта наблюдения в поле зрения при поворотах головы или вращении тела.
Сенсорные (корковые) реакции возникают при определенной интенсивности раздражений и выражаются чувством головокружения, разнообразными пространственными иллюзиями собственного положения или окружающих предметов относительно друг друга.
Вестибулярно-вегетативные реакции в нормальных условиях обеспечивают необходимый в данный момент времени уровень обменных процессов в мышечной системе. При длительном воздействии вестиб. раздражений, особенно ускорений, противоположных по направлению, у некоторых людей возможен срыв адаптационных механизмов вегетативной нервной системы с возникновением патологических реакций в виде: тошноты, рвоты, саливации, потливости, головокружения, тахи- или брадикардии и др. Эти состояния получили название кинетозов — болезни передвижения (морская, воздушная, космическая, транспортная). У новорожденных и больных с удаленными лабиринтами кинетозов не наблюдается.
Дата: 2019-07-30, просмотров: 926.