На обратной транскрипции основано размножение ретровирусов (вирусы, у которых геномом служит не ДНК, как обычно, а РНК) и ретротранспозонов (являются транспозиционными элементами, которые не имеют вирионных частиц, и, следовательно, в отличие от ретровирусов, не могут независимо «переносить себя» между клетками), образование так называемых ретропсевдогенов (или процессированные псевдогены это ретропоследовательности, которые потеряли свою функцию, они несут все признаки функциональных ретропоследовательностей, но имеют молекулярные дефекты, которые не дают им экспрессироваться) и достройка кончиков хромосом (теломер), укорачивающихся при каждом клеточном делении. Если молекула ДНК повреждена -- например, подверглась разрыву (double-strand break, DSB) -- для ее починки необходима матрица, в которой последовательность нуклеотидов соответствует исходному, «правильному» состоянию поврежденного участка. Ранее считалось, что в качестве таких матриц всегда используются другие молекулы ДНК. Позже было установлено, что иногда эти ДНК-матрицы синтезируются путем обратной транскрипции на основе РНК при участии ретротранспозонов.
Этапы обратной транскрипции:
1. Наращивание тРНК-праймера на матрицах U5 и R в направлении 3'>5'. Роль РНК-праймера выполняет одна из клеточных тРНК (например, триптофановая, пролиновая и т.д.). На расстоянии примерно 100 -- 200 нуклеотидов от 5'-конца РНК (для каждого вируса -- это величина постоянная) имеется участок, комплементарный З'-концевой последовательности молекулы тРНК, который используется в качестве затравки. Этот участок обычно обозначают как pbs (от англ.primer binding site участок связывания затравки). Обратная транскриптаза синтезирует сегмент ДНК, комплементарный 5'-концевой последовательности вирусной РНК. Этот сегмент принято называть (-) «strong-stop» ДНК, поскольку синтез ДНК после завершения копирования 5'-конца матрицы временно останавливается. (-) «strong-stop» ДНК содержит последовательности, комплементарные концевому району R и району U5. Таким образом, синтез ДНК начинается недалеко от 5'-конца матрицы и образуется короткий продукт. Но этот короткий продукт (-) «strong-stop» имеет последовательность, комплементарную также и З'-концу вирусной РНК, а как известно, для снятия ДНК-копии с З'-конца матрицы всегда требуется праймер. У ретровирусов комплемент З'-конца матрицы производится в «удобном» месте, а затем переносится на «свое» место. Это происходит следующим образом: 5'-конец вирусной РНК, образующий дуплекс с (-) «strong-stop» ДНК, разрушается под влиянием присущей обратной транскриптазе активности РНКазы Н.
2. РНКаза Н, специфичная к РНК в составе гибридного РНК-ДНК дуплекса, расщепляет сегмент РНК этого дуплекса. В результате (-) «strong-stop» (RU5) оказывается в однонитевой форме и может взаимодействовать с З'-концом (с участком R) той же самой или другой молекулы вирусной РНК, поскольку на З'-конце РНК имеется повтор R..
3. Новосинтезированная короткая цепь ДНК вместе с праймером «перепрыгивает» на З'-конец матрицы и взаимодействует там с комплементарным ей участком К.
4. Цепь ДНК удлиняется, в качестве матрицы используется остальная часть вирусной РНК. На этой стадии в качестве затравки выступает уже (-)«strong-stop» ДНК; элонгация затравки приводит к синтезу (-) цепи ДНК, в которой отсутствует комплемент района RU5, поскольку соответствующий участок (+) матрицы был разрушен РНКазой Н.
5. К моменту завершения синтеза первой цепи ДНК большая часть вирусной РНК разрушается РНКазой Н.
6. Синтез З'-конца второй цепи ДНК.
7. Удаление тРНК и оставшегося участка вирусной (+) РНК РНКазой Н.
8. Второй прыжок, в результате которого новосинтезированная вторая цепь ДНК комплементарно взаимодействует с тРНК-связывающей последовательностью первой цепи.
9. Удлинение З'-концов каждой цепи, образование дуплекса ДНК.
Вся последовательность реакций протекает без явного участия ферментов репликации клетки-хозяина (топоизомеразы, хеликазы, праймазы, ДНК-связывающего белка, лигазы и т.д.). При этом следует отметить, что молекулы вирусных ДНК длиннее молекул вирусных РНК, которые послужили матрицей для обратной транскрипции. Действительно, к 5'-концу (+) цепи вирусной ДНК добавилась последовательность U3, а к 3 -концу этой цепи -- последовательность U5. В результате на концах молекулы вирус специфической ДНК появился длинный (несколько сотен нуклеотидов) концевой повтор (ДКП или LTR.), имеющий структуру U3U5.
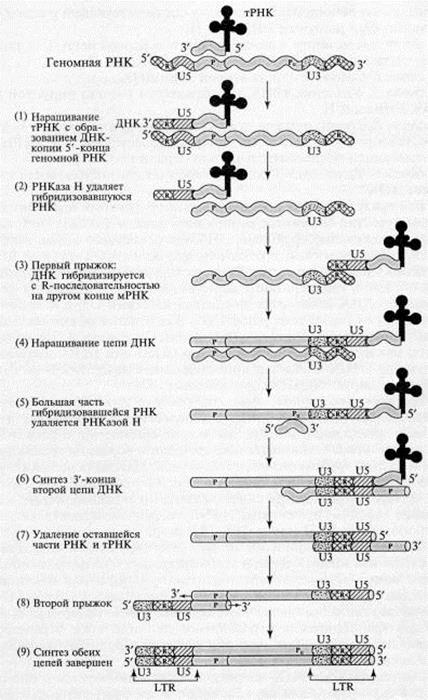
Схема обратной транскрипции ретровирусной РНК с образованием двуцепочечной ДНК (Darnell J., et.al. Molecular Cell Biology. - N. Y.: Scientific Amer. Books, 1986. - P. 1052
Мы все знаем что такое ПЦР МОЛОДЦЫ!!! А я забыл…
Буфер-муфер, и не только, все здесь --- https://biomolecula.ru/articles/metody-v-kartinkakh-polimeraznaia-tsepnaia-reaktsiia Ниже все таки привел ход реакции! Но ты должен знать что там есть вообще!
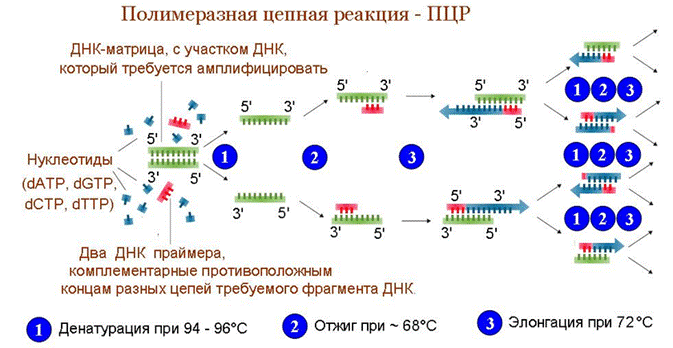
Ход реакции ПЦР
Обычно при проведении ПЦР выполняется 20—35 циклов, каждый из которых состоит из трёх стадий.
Денатурация
Двухцепочечную ДНК-матрицу нагревают до 94—96 °C (или до 98 °C, если используется особенно термостабильная полимераза) на 0,5—2 мин, чтобы цепи ДНК разошлись. Эта стадия называется плавлением (денатурацией ), так как разрушаются водородные связи между двумя цепями ДНК. Обычно перед первым циклом проводят длительный прогрев реакционной смеси в течение 2—5 мин для полной денатурации матрицы и праймеров.
Отжиг
Когда цепи разошлись, температуру понижают, чтобы праймеры могли связаться с одноцепочечной матрицей. Эта стадия называется отжигом. Температура отжига зависит от состава праймеров и обычно выбирается на 5 градусов меньше, чем температура плавления праймеров. Неправильный выбор температуры отжига приводит либо к плохому связыванию праймеров с матрицей (при завышенной температуре), либо к связыванию в неверном месте и появлению неспецифических продуктов (при заниженной температуре). Время стадии отжига — 30 сек, одновременно, за это время полимераза уже успевает синтезировать несколько сотен нуклеотидов. Поэтому рекомендуется подбирать праймеры с температурой плавления выше 60 °C и проводить отжиг и элонгацию одновременно, при 60—72 °C.
Элонгация
ДНК-полимераза реплицирует матричную цепь, используя праймер в качестве затравки. Это — стадия элонгации. Полимераза начинает синтез второй цепи от 3'-конца праймера, который связался с матрицей, и движется вдоль матрицы, синтезируя новую цепь в направлении от 5'- к 3'-концу. Температура элонгации зависит от полимеразы. Часто используемые полимеразы Taq и Pfu наиболее активны при 72 °C. Время элонгации зависит как от типа ДНК-полимеразы, так и от длины амплифицируемого фрагмента. Обычно время элонгации принимают равным одной минуте на каждую тысячу пар оснований. После окончания всех циклов часто проводят дополнительную стадию финальной элонгации, чтобы достроить все одноцепочечные фрагменты. Эта стадия длится 7—10 мин.
Количество специфического продукта реакции (ограниченного праймерами) теоретически возрастает пропорционально 2n — 2n, где n — число циклов реакции[17]. На самом деле эффективность каждого цикла может быть меньше 100 %, поэтому в действительности P ~ (1+E)n, где P — количество продукта, Е — средняя эффективность цикла.
Число «длинных» копий ДНК тоже растет, но линейно, поэтому в продуктах реакции доминирует специфический фрагмент.
Рост требуемого продукта в геометрической прогрессии ограничен количеством реагентов, присутствием ингибиторов, образованием побочных продуктов. На последних циклах реакции рост замедляется, это называют «эффектом плато».
Дата: 2019-05-28, просмотров: 448.