Насекомые
M многообразие насекомых и их значение
Роль насекомых в природе обусловлена их численностью и разнообразием. Растительноядные насекомые питаются растениями, и тем самым регулируют их растительный прирост, поедая основную часть. А вот паразитические и хищные насекомые считаются регуляторами численности представителей животных, которыми они же и питаются. Таким образом, большое значение насекомые имеют как потребители животных и растительных остатков.
Насекомые – это опылители растений, поэтому играют роль в их размножении. Также они берут участие в почвообразовании. Эти организмы не только рыхлят почву, но и обогащают ее перегноем. Насекомые одновременно выполняют роль санитаров и являются участниками круговорота веществ в природе. К тому же они считаются важнейшим элементом пищевых пирамид: многие животные питаются ими (земноводные, рыбы, пресмыкающиеся, млекопитающие, беспозвоночные и птицы). Значение насекомых в жизни человека Человек одомашнил насекомых для своей выгоды. Медоносная пчела производит воск, мед, прополис, маточное молочко и апилак. На основе этих «даров» люди развили пчеловодство. Одомашненный тутовый шелкопряд снабжает нас шелковой нитью. Также ценную продукцию дают — лаковые червецы (их воскоподобное вещество используется в электро- и радиотехнике), гусеницы дубового коконопряда (из его шелковой нити изготовляют ткань чесуча), карминные червецы (продуцируют красную краску — кармин), жуки-нарывники (выделяют кантаридин, из которого изготавливают нарывной пластырь). Многие насекомые-опылители повышают урожаи ягод, семян, плодов культурных растений — овощных, цветочных, плодово-ягодных, кормовых. Хищники и паразиты помогают уничтожить вредителей сельского хозяйства. Вред насекомых в жизни человека Отрицательные значения насекомых в том, что они являются переносчиками опасных возбудителей разных заболеваний. На своих лапках насекомые переносят грибковые бактерии, микробы и другие вредные микроорганизмы, которые способствуют гниению овощей, фруктов. Также они отлаживают личинки в продуктах питания и даже в ранах самого человека.
Физиология рыб.
Сенсорные системы рыб.
Внешние стимулы, воспринимаемые представителями класса рыб:
· Электромагнитная и тепловая энергия
· Свет
· Тепло/холод
· Электричество
· Магнитная энергия
· Механическая энергия
· Звук/вибрация
· Давление/осмотическое давление
· Гравитация
· Прикосновение
· Инерция
· Химические факторы
· Влажность
· Вкус
· Запах
У рыб контакт с внешней средой еще более плотный, чем у высших позвоночных. Поэтому и контроль за изменениями во внешней среде у рыб должен быть более чутким. Этому способствует хорошо развитый рецепторный аппарат. Рыбы реагируют на видимый человеком свет, электромагнитные поля.
Для восприятия и анализа наиболее важных сведений из внешней среды эволюция снабдила животных высокоспециализированными структурами - сенсорными системами, которые обладают высокой чувствительностью и избирательной реактивности по отношению к свету, звуку, химическому составу и температуре окружающей среды, электромагнитному полю, изменению гравитации, давления,
Сенсорная система включает в себя рецепторный аппарат (глаз, ухо, ампулы Лоренцини и др.) и анализирующий аппарат в составе центральной нервной системы.
Обращает на себя внимание то, что сенсорные органы у рыб не так четко дифференцированы по функциям, как у высших позвоночных. Например, у рыб трудно назвать орган слуха. Экспериментально установлено, что рыбы реагируют на звук. Но за восприятие колебаний воды у рыбы отвечают несколько органов: боковая линия, лабиринт, плавательный пузырь, а у пластинчатожаберных еще и особые образования - ампулы Лоренцини на голове и окончаниях лицевого нерва.
Более того, термин "органы чувств" применительно к рыбам зачастую лишен первоначального смысла, так как сенсорная информация может и не поступать в центральную нервную систему. В этом случае она не подвергается чувственной оценке, следовательно, и структуры, отвечающие за эту рецепцию факторов внешней среды, нельзя называть органами чувств.
Тем не менее, сенсорные системы рыб обеспечивают надежную связь между водной средой и организмом рыбы. Тот факт, что при отсутствии больших полушарий мозга (и тем более кортикальных структур) рыбы проявляют эмоциональные реакции на действие раздражителей из внешней среды, за формирование которых у рыб отвечает лимбическая система, оправдывает применение термина "сенсорные" (чувственные) системы при изучении физиологии рыб
Перенос газов кровью.
Помимо жабр у рыб в газообмене принимают участие кожа, желудочно-кишечный тракт, плавательный пузырь, специальные органы. Жабры. Основная нагрузка в обеспечении организма рыб кислородом и удалении из него углекислого газа ложится на жабры. Жабры хорошо приспособлены к газообмену в водной среде. Кислород переходит в капиллярное русло жабр по градиенту парциального давления, который у рыб составляет 40-100 мм рт. ст. Такова же причина перехода кислорода из крови в межклеточную жидкость в тканях. Жаберная поверхность в 10-60 раз превышает площадь тела рыбы. К тому же жабры, высокоспециализированные на газообмене органы. Самое совершенное строение жаберного аппарата характерно для костистых рыб. Основой жаберного аппарата являются 4 пары жаберных дуг. На жаберных дугах располагаются хорошо васкуляризированные жаберные лепестки, образующие дыхательную поверхность.По стороне жаберной дуги, обращенной в ротовую полость, располагаются более мелкие структуры - жаберные тычинки, которые в большей мере отвечают за механическую очистку воды по мере ее поступления из ротовой полости к жаберным лепесткам.Поперечно жаберным лепесткам располагаются микроскопические жаберные лепесточки, которые и являются структурными элементами жабр как органов дыхания. Покрывающий лепесточки эпителий имеет клетки трех типов: респираторные, слизистые и опорные. Площадь вторичных ламелей и, следовательно, дыхательного эпителия зависит от биологических особенностей рыбы - образа жизни, интенсивности основного обмена, потребности в кислороде.Однако известно, что крупные и активные виды, например тунец, рта не закрывают, и дыхательные движения жаберных крышек у них отсутствуют. Такой тип вентиляции жабр называют "таранным"; он возможен только при больших скоростях перемещения в воде.Для прохождения воды через жабры и движения крови по сосудам жаберного аппарата характерен противоточный механизм, обеспечивающий очень высокую эффективность газообмена. Пройдя через жабры, вода теряет до 90 % растворенного в ней кислорода. Жаберный газообмен может быть эффективным только при постоянном токе воды через жаберный аппарат. Жаберное дыхание следует рассматривать как очень эффективный механизм газообмена в водной среде с точки зрения эффективности извлечения кислорода, а также энергозатрат на этот процесс. В том случае, когда жаберный механизм не справляется с задачей адекватного газообмена, включаются другие (вспомогательные) механизмы.
Кожное дыхание. Кожное дыхание имеет существенное значение для видов, ведущих малоподвижный образ жизни в условиях низкого содержания кислорода, или, вообще, покидающих кратковременно водоем (угорь, илистый прыгун, сомы). Изучение онтогенетического развития рыб свидетельствует о том, что кожное дыхание первично по отношению к жаберному. Эмбрионы и личинки рыб осуществляют газообмен с окружающей средой через покровные ткани. Интенсивность кожного дыхания усиливается с повышением температуры воды, так как повышение температуры усиливает обмен веществ и снижает растворимость кислорода в воде.В целом, интенсивность кожного газообмена определяется морфологией кожи. Механизм кожного дыхания у животных изучен явно недостаточно. Важную роль в этом процессе играет кожная слизь, в составе которой обнаруживается и гемоглобин, и фермент карбоангидраза.
Кишечное дыхание. В экстремальных условиях (гипоксия) кишечное дыхание используется многими видами рыб. У таких рыб (сомик, пескарь) воздух заглатывается и перистальтическими движениями кишечника направляется в специализированный отдел. Заглоченный пузырек атмосферного воздуха в кишке находится под определенным давлением, что повышает коэффициент диффузии кислорода в кровь. В этом месте кишка обеспечивается венозной кровью, поэтому возникают хорошая разница парциального давления кислорода и углекислого газа и однонаправленность их диффузии. Плавательный пузырь не только обеспечивает рыбе нейтральную плавучесть, но и играет определенную роль в газообмене. Он бывает открытым (лососевые) и закрытым (карп). Открытый пузырь связан воздушным протоком с пищеводом, и его газовый состав может быстро обновляться. В закрытом пузыре изменение газового состава происходит только через кровь.В стенке плавательного пузыря имеется особая капиллярная система, которую принято называть "газовой железой". Капилляры железы образуют круто изогнутые противоточные петли. Однако противоточный механизм кровотока в газовой железе приводит к тому, что этот кислород плазмы диффундирует в полость пузыря. Таким образом, пузырь создает запас кислорода, который используется организмом рыбы в неблагоприятных условиях.Другие приспособления для газообмена представлены лабиринтом (гурами, лялиус, петушок), наджаберным органом (рисовый угорь), легкими (двоякодышащие), ротовым аппаратом (окунь ползун), глоточными полостями (Ophiocephalussp.). Морфологически и функционально с органами дыхания связаны псевдобранхии - особые образования жаберного аппарата. То, что к этим структурам притекает кровь от жабр, насыщенная кислородом, свидетельствует о том.что они не участвуют в обмене кислорода. Однако наличие большого количества карбоангидразы на мембранах псевдобранхии допускает участие этих структур в регуляции обмена углекислого газа в пределах жаберного аппарата. Функционально с псевдобранхиями связана так называемая сосудистая железа, расположенная на задней стенке глазного яблока и окружающая зрительный нерв. Сосудистая железа имеет сеть капилляров, напоминающую таковую в газовой железе плавательного пузыря. Есть точка зрения, что сосудистая железа обеспечивает снабжение сильно насыщенной кислородом кровью сетчатки глаза при максимально низком поступлении в нее углекислого газа.
Перенос газов кровью. Основным транспортировщиком кислорода в крови у рыб выступает гемоглобин. Гемоглобин рыб функционально делится на два типа- чувствительный к кислоте и нечувствительный к кислоте. Чувствительный к кислоте гемоглобин при понижении рН крови утрачивает способность связывать кислород.Нечувствительный к кислоте гемоглобин не реагирует на величину рН. У некоторых арктических и антарктических видов рыб гемоглобина в крови нет вообще. В определенных условиях с транспортированием газов справляется одна плазма. Однако в обычных условиях у подавляющего большинства рыб газообмен без гемоглобина практически исключен. Диффузия кислорода из воды в кровь протекает по градиенту концентрации. Градиент сохраняется, когда растворенный в плазме кислород связывается гемоглобином, т.е. диффузия кислорода из воды идет до полного насыщения гемоглобина кислородом. Транспорт углекислого газа кровью осуществляется по-другому. Роль гемоглобина в переносе углекислого газа в виде карбогемоглобина невелика. Гемоглобин переносит не более 15 % углекислого газа. Основной транспортной системой для переноса углекислого газа является плазма крови. Попадая в кровь в результате диффузии из клеток, углекислый газ вследствие его ограниченной растворимости создает повышенное парциальное давление в плазме и таким образом должен тормозить переход газа из клеток в кровяное русло. На самом деле этого не происходит. В плазме под влиянием карбоангидразы эритроцитов осуществляется реакция СО2 + Н2О->Н2СО3-->Н+ + НСО3
Образующийся бикарбонат с кровью поступает в жаберный эпителий, который также содержит карбоангидразу. Поэтому в жабрах происходит преобразование бикарбонатов в углекислый газ и воду. Далее по градиенту концентрации СО2 из крови диффундирует в омывающую жабры воду.
Физиология птицы.
37.
Стадии формирования яйца
· формирование яйца состоит из двух различных процессов:
· рост и созревание зародышевой клетки;
· отложение органического вещества — желтка,,белканоболочек яйца.
· Органами, где образуется яйцо, являются яичник и яйцевод.
В яйцеводе яйцо последовательно приобретает белок, две подскорлулные оболочки и скорлупу.
Сначала яйцо существует в форме овоцита, находящегося в яичнике курицы. Этот овоцит - зачаток яйца - впоследствии станет желтком. Образование самого яйца происходит в яйцеводе.
Зачатки будущих яиц закладываются еще в эмбриональный период. К середине инкубации куриный эмбрион уже имеет полный набор микроскопических по размеру яйцеклеток (примерно от 600 до 3600 шт.), находящихся в яичнике. После вылупления птенца вокруг каждой яйцеклетки образуется оболочка — фолликул, продуцирующий желтковую массу. Яйцеклетки растут и к 3 х-недельному возрасту цыпленка достигают диаметра примерно 0,05 мм, а к началу полового созревания увеличиваются до 1 мм. Перед яйцекладкой часть яйцеклеток вступает в фазу бурного роста. Освободившись от фолликула, овоцит попадает в воронку — начало яйцевода, а затем в яйцевод, где происходит его оплодотворение и дальнейшее развитие. Яйцевод состоит из 5 отделов, каждый из которых участвует в образовании определенных частей яйца. Воронка яйцевода, расположенная около яичника, принимает в себя зрелый желток после овуляции. Здесь происходит оплодотворение яйцеклетки, а затем образуется густой волокнистый слой белка, часть которого в дальнейшем при винтообразном движении желтка скручивается в виде спиральных полупрозрачных образований — халаз, или градинок, они поддерживают желток в центре яйца. В воронке яйцевода желток задерживается до 20 мин, а затем через узкую ее часть (шейку) поступает в самый длинный белковый отдел, в котором стенками яйцевода вырабатывается альбумин, из которого состоит белок яйца, этот процесс длится примерно четыре час. Яйцо, снабжённое теперь белком, продолжает свой путь и поступает в перешеек яйцевода, в котором выделяется клейкое нитевидное вещество, которое, переплетаясь пучками, образует две подскорлупные оболочки. Сначала они прочно соединены, затем разделяются. Яйцо пребывает в перешейке немногим более 1 ч. Перешеек без четко выраженной границы переходит в самую широкую часть яйцевода — матку, к этому моменту оболочки ослабляются и как бы «плавают» вокруг белка. В матке формируется скорлупа. В течение последующих пяти часов происходит постепенное проникновение воды и солей сквозь оболочки до тех пор, пока они снова не растянутся и яйцо «вздуется». Этот предварительный процесс необходим для кальцинирования скорлупы, которое происходит в течение последующих 15—16 часов. Через 3−4 ч после начала образования скорлупы последняя представляет собой тонкую, хрупкую, легкосминаемую оболочку. В матке яйцо находится в течение 16 — 20 ч.
Скорлупа состоит главным образом из кальцита — кристаллов карбоната кальция. Она трехслойная: внутренний слой, составляющий треть толщины скорлупы, состоит из конических кристаллов, скреплённых белковыми мостиками, следующий слой состоит из тесно прижатых друг к другу столбчатых кристаллов карбоната кальция. Всё это покрыто тонкой кожицей (кутикулой), которая образуется непосредственно перед откладыванием яйца. В последние часы формирования яйца в матке скорлупа окрашивается за счет пигмента переработанных печенью эритроцитов и покрывается надскорлупной пленкой, или кутикулой, состоящей из тонкого слоя слизи. Полностью сформированное яйцо через последний отдел яйцевода — влагалище — выталкивается наружу. При этом матка сильно выпячивается, перекрывая отверстие клоаки, благодаря чему яйцо при снесении не загрязняется.
У кур яйцо по яйцеводу проходит обычно в течение 23 — 26 ч. Из них примерно 80 % времени оно находится в матке.После снесения и остывания яйца желток и белок слегка уменьшаются в объеме, на тупом полюсе подскорлупные оболочки расходятся, и между ними образуется воздушная камера. Воздушная камера сразу же после остывания яйца имеет диаметр менее 1 см, затем она увеличивается в зависимости от сроков хранения, температуры и влажности окружающего воздуха. Размер воздушной камеры может являться показателем свежести яиц.
39-41.
Особенности дыхания.
Дыхание представляет совокупность процессов, обеспечивающих потребление организмом кислорода и выделение углекислого газа. Оно включает следующие процессы:
1) обмен воздуха между внешней средой и альвеолами легких (внешнее дыхание),
2) обмен газами между альвеолярным воздухом и кровью (газообмен в легких),
3) перенос (транспорт) газов кровью,
4) обмен газов между кровью и тканями,
5) потребление кислорода клетками и выделение ими углекислоты.
Внешнее дыхание осуществляется в результате ритмических дыхательных движений грудной клетки - вдоха и выдоха. При вдохе объем грудной клетки увеличивается, легкие расширяются, давление в них становится ниже атмосферного и воздух через воздухоносные пути поступает в легочные альвеолы.
Различают три типа дыхания: грудной, брюшной и смешанным При грудном, или реберном, типе в дыхании принимает участив только стенка грудной клетки, при брюшном, или диафрагмальном, дыхательные движения совершаются мышцами живота и диафрагмой. У сельскохозяйственных животных обычно наблюдается смешанный тип дыхания, при котором принимают участие как грудные, так и брюшные мышцы.
Координированную ритмичную деятельность органов дыхания обеспечивает дыхательный центр, расположенный в продолговато том мозгу. Возбуждение дыхательного центра вызывается как рефлекторно импульсами, которые поступают к нему из легких, рецепторов аорты, синусов сонной артерии и самых разнообразных экстеро- и интерорецепторов, а также гуморально - химическими веществами (углекислотой и др.), приносимыми к нему кровью. Кроме того, возбуждение дыхательного центра может вызываться автоматически продуктами обмена веществ, которые вырабатываются в самом центре.
При характеристике объемов легочного воздуха различают дыхательный, дополнительный, резервный и остаточный воздух, а также жизненную емкость легких.
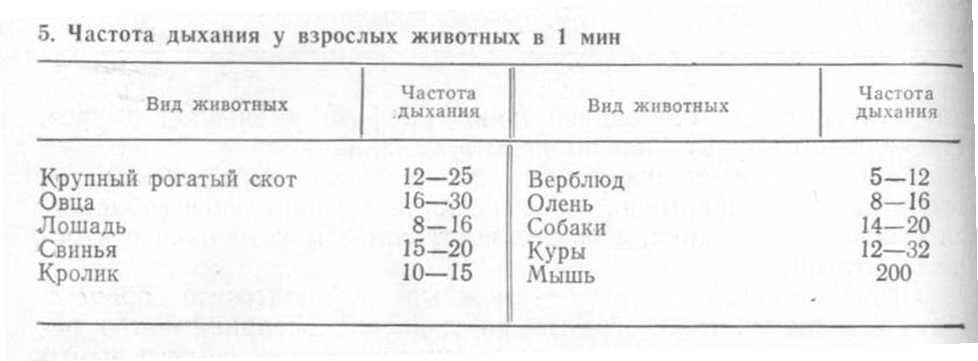
Дыхательный воздух - это объем воздуха, который животное вдыхает и выдыхает в спокойном состоянии. Он у крупного рогатого скота и лошади составляет около 6 л. Дополнительный воздух- это тот воздух, который животное может вдохнуть при глубоком вдохе сверх дыхательного воздуха. У крупных животных он равняется примерно 12 л. После нормального спокойного выдоха животное может дополнительно выдохнуть около 12 л воздуха - это резервный воздух.
Дыхательный, дополнительный и резервный воздух составляет жизненную емкость легких, которая у крупного рогатого скота и лошадей равна около 30 л.
Однако даже после максимально глубокого выдоха легкие не освобождаются полностью от воздуха, в них остается остаточный воздух, объем которого у лошади примерно составляет 10 л.
Сопоставление состава вдыхаемого и выдыхаемого воздуха показывает, что около 4,5% кислорода из вдыхаемого воздуха остается в организме и около 4% углекислого газа выделяется из него с выдыхаемым воздухом (табл. 6).
Координированную ритмичную деятельность органов дыхания обеспечивает дыхательный центр, расположенный в продолговато том мозгу. Возбуждение дыхательного центра вызывается как рефлекторно импульсами, которые поступают к нему из легких, рецепторов аорты, синусов сонной артерии и самых разнообразных экстеро- и интерорецепторов, а также гуморально - химическими веществами (углекислотой и др.), приносимыми к нему кровью. Кроме того, возбуждение дыхательного центра может вызываться автоматически продуктами обмена веществ, которые вырабатываются в самом центре.
У птиц имеется ряд характерных морфологических и функциональных отличий дыхания по сравнению с млекопитающими животными. При вдохе воздух поступает в легкие и далее по мелким бронхам в воздухоносные мешки. При выдохе грудная клетка сжимается и воздух из воздухоносных мешков проходит через легкие в обратном направлении. Следовательно, через альвеолы воздух проходит как во время вдоха, так и выдоха, дважды отдавая кислород в кровь.
Таким образом, воздухоносные мешки (шейные, межключичные, передне- и заднегрудные, брюшные) создают резерв воздуха. Газообмен в них не происходит. Воздух, заполняющий воздухоносные мешки, охлаждает тело птицы. Воздухоносные мешки, кроме того, облегчают полет птиц, а у водоплавающих птиц - передвижение их по воде.
Жизненная емкость легких у кур составляет 13, а у уток - 20 мл, емкость воздухоносных мешков - соответственно 169 и 315 мл. Частота дыхательных движений в минуту составляет у кур 12-32, уток - 15-18, гусей - 9-10 и у индеек- 15-20.
Газообмен в легких и механизм регуляции дыхания у птиц и млекопитающих аналогичны.
Ротовое пищеварение
Передний отдел пищеварительного аппарата - ротовая полость - является начальным отделом пищеварительной трубки, куда при естественных условиях, прежде всего, поступает пища и где она подвергается первоначальной механической и химической обработке.
Пища находится в ротовой полости сравнительно короткое время тем не менее, достаточное для того, чтобы она могла быть размельчена и смочена слюной, для формирования пищевого комка, т.е. подготовлена к проглатыванию. Пищеварение во рту сводится главным образом к механической обработке пищи. Химическое же воздействие слюны на пищевые вещества (углеводы) весьма ничтожно из-за непродолжительного пребывания пищевой массы в ротовой полости.
Ротовое пищеварение включает три этапа: прием корма, собственно ротовое пищеварение - жевание и ослюнение, акт глотания.
Прием корма
Пищеварению предшествует акт приема корма. В нем участвуют следующие исполнительные органы: губы, язык, жевательные мышцы, челюсти, зубы, слюнные железы, глотка, пищевод, желудок. В этом процессе принимают участие кроме органов пищеварения органы движения. Система обеспечивает поиск и прием корма, удовлетворение потребностей организма в питательных, минеральных веществах, витаминах и воде.
Поиск корма
Поиск корма обусловлен чувством голода и связан со зрительной, обонятельной, вкусовой рецепциями, осязанием его языком и губами. Наступающее после кормления состояние насыщения определяется двумя факторами: сенсорным и метаболическим насыщением.
Сенсорное насыщение связано с влиянием корма на механо- и хеморецепторы полости рта и желудка, в результате прием корма заканчивается до начала процесса всасывания.
Метаболическое наступает через 1,5-2 ч после приема корма. Причина его - повышение уровня доступной глюкозы и аминокислот при всасывании жиров. У животных при свободном доступе к корму пищеварительный тракт практически не бывает пустым.
Жевание
Захваченная животным, порция корма направляется на поверхность зубов и пережевывается.
Жевание - сложный рефлекторный акт, состоящий из последовательных сокращений жевательной мускулатуры.
Жевание имеет важное значение для последующего хода пищеварения. Оно способствует оценке вкусовых качеств пищи, стимулирует отделение пищеварительных соков, которые быстрее и лучше проникают в размельченную пищу, а это обуславливает более полное переваривание и последующее ее всасывание.
Число жевательных движений при пережевывании грубых кормов у крупного рогатого скота более 15 тыс., у овец-12 тыс. Крупный рогатый скот корм захватывает языком, направляет на резцы нижней челюсти, сдавливает между резцами нижней челюсти и зубной пластиной резцовой кости и рывком головы отрывает захваченную часть. Концентраты, корнеплоды и клубнеплоды из кормушки захватываются также губами и зубами. Захваченная порция корма пережевывается. Животное совершает 22…30 жевательных движений в зависимости от объема, структуры и состава порции корма. При приеме грубых кормов число жевательных движений больше. Время пережевывания одной порции составляет 20…50 с.
Тщательно пережевывают корм свиньи. Прием корма у них сопровождается чавканьем вследствие выхода воздуха, попадающего при жевании в ротовую полость через открытые щели рта.
Лошади пережевывают корм длительно и тщательно. У них отмечают функциональную асимметрию жевания: жуют на одной стороне челюстей, затем через 30…60 мин меняют сторону жевания. Лошадь пережевывает 1 кг овса за 8…9 мин; на формирование пищевого кома затрачивает около 50 жевательных движений.
Кролик ест понемногу, но часто. Взрослый кролик потребляет пищу 25-30 раз в сутки, причем большее число раз - ночью и рано утром, а молодняк примерно в два раза чаще. Кролик - грызун, две трети его рациона должны состоять из кормов, богатых клетчаткой.
У жвачных животных после приема корма происходит отрыгивание и повторное пережевывание принятого корма - жвачка.
Жвачка - приспособительная реакция у жвачных животных, проявляющаяся в отрыгивании поверхностно пережеванных порций корма из рубца, повторном пережевывании и проглатывании их, что обеспечивает дальнейшее переваривание принимаемых растительных кормов. Жвачка осуществляется периодами; в течение суток 6…8 периодов по 14…50 мин. Каждый жвачный период состоит из отдельных жвачных циклов: 8…40 циклов в периоде. Каждый цикл состоит из трех фаз: отрыгивание порции корма из пищеводной области рубца через пищевод обратно в ротовую полость, пережевывание этой порции, проглатывание повторно пережеванной порции корма. Крупный рогатый скот совершает при пережевывании каждой порции отрыгнутого корма 40…60 жевательных движений, овцы - 50…60
Слюноотделение
Корм в ротовой полости смешивается со слюной, которая вырабатывается околоушной, подъязычной и подчелюстной железами, я также мелкими железами слизистой оболочки ротовой полости.
Слюна - бесцветная, слегка опалеоцирующая жидкость слабощелочной реакции: рН слюны у жвачных животных - около 8,0, у свиней - 7,32, у лошади - 7,5. Она состоит из 99-99,4% воды и 0,6-1% сухих веществ. Из органических веществ в слюне имеются хлориды, сульфаты, карбонаты кальция, натрия, калия, магния, а также мочевина, мочевая кислота, муцин, который обусловливает ее вязкость. Слюна содержит вещества, обладающие антимикробным действием - лизоцим и ингибан и два фермента - амилазу, расщепляющую крахмал до мальтозы и декстринов, и глюкозидазу (мальтазу), превращающую мальтозу в глюкозу.
Слюноотделение регулируется центральной нервной системой и представляет собой сложную рефлекторную реакцию. Слюноотделительные рефлексы могут быть условными и безусловными. При безусловном рефлексе корм раздражает рецепторы ротовой полости, по центростремительным (афферентным) нервам возбуждение передается в пищевой центр продолговатого мозга, а оттуда по центробежным симпатическим и парасимпатическим нервам импульсы посылаются к слюнным железам, и они начинают выделять слюну. Условно-рефлекторное слюноотделение появляется в ответ на раздражение зрительных, слуховых и обонятельных рецепторов, т.е. на вид и запах корма. Если кормление животного сопровождать несколько раз действием условного раздражителя (звук, свет, обстановка опыта и др.), то через некоторое время отделение слюны будет происходить при действии одного условного рефлекса. В основе условного слюноотделительного рефлекса лежит образование в коре головного мозга временной связи между центрами условного и безусловного рефлексов.
Слюноотделение у лошадей
Слюноотделение у животных разных видов несколько отличается. Суточное кол-во слюны у взрослой лошади достигает 40-50 литров. Количество секрета увеличивается при скармливании раздробленных кормов (плющенное зерно, сечка), добавлением соли. При недостаточном поении количество слюны уменьшается наполовину. Еще одна особенность - слюна у лошади выделяется только при приеме корма. Основная ее роль смачивание пищи, хотя в ней обнаружено небольшое количество амилолитических (расщепляющих углеводы) ферментов. Но основная роль слюны - физическая подготовка корма к перевариванию. Большое значение слюна лошади имеет в желудочном пищеварении, так как она создает в желудке щелочную среду, необходимую для действия ферментов растительных кормов и микрофлоры.
Слюноотделение у свиней
У свиней слюны может выделяться (в зависимости от корма) от 5 до 15 л в сутки. В слюне находится фермент птиалин, или амилаза, расщепляющий крахмал до сахара. Слюноотделение больше всего возбуждается грубыми и сухими кормами, меньше всего - зелеными и сочными. Обладает выраженной ферментативной активностью.
Глотание
Перемещение корма из ротовой полости в желудок - глотание. При этом пищевой ком прижимается к нёбу языком и проталкивается в глотку. В это время нёбная занавеска поднимается и закрывает носоглотку, а корень языка прижимает надгортанник к гортани, прикрывая путь в верхние дыхательные пути. Отверстие пищевода при этом расширяется и подтягивается к глотке, сильным сокращением мышц глотки корм проталкивается в пищевод. В акте глотания выделяют произвольную фазу, когда пищевой ком располагается в ротовой полости до корня языка и животное еще может выбросить его, а далее уже наступает непроизвольная фаза, когда осуществляются глотательные движения. Центр глотания связан с другими центрами продолговатого мозга, поэтому в момент глотания тормозится дыхательный центр, в результате чего происходит задержка дыхания и учащение работы сердца. Высшие центры глотания расположены в гипоталамической части промежуточного и в коре головного мозга. Глотание при отсутствии в ротовой полости корма или слюны практически трудно осуществимо или невозможно.
Состав и функции слюны
Смотри вопрос 59.
Пищеварение в желудке
Желудочное пищеварение
Из пищевода корм поступает в желудок, где задерживается определенное время и подвергается дальнейшей механической и химической обработке.
Настоящее пищеварение принятого корма начинается в желудке. Пищеварение в желудке связано с секреторной деятельностью желудочных желез, двигательной активностью мышц желудка и деятельностью кардиального и пилорического сфинктеров его.
По характеру пищеварения сельскохозяйственных животных подразделяют на два типа. Первый тип - животные с однокамерным желудком (лошади, свиньи, собаки). У них желудочно-кишечное пищеварение; переваривание корма в основном происходит под влиянием пищеварительных ферментов, а продукты переваривания всасываются в кишечнике. Второй тип - жвачные животные (крупный рогатый скот, овцы, козы, буйволы, верблюды, северные олени и др.). У них преобладает желудочное пищеварение, и значительная часть корма переваривается без участия ферментов пищеварительных соков под влиянием ферментов микроорганизмов, населяющих пищеварительный тракт.
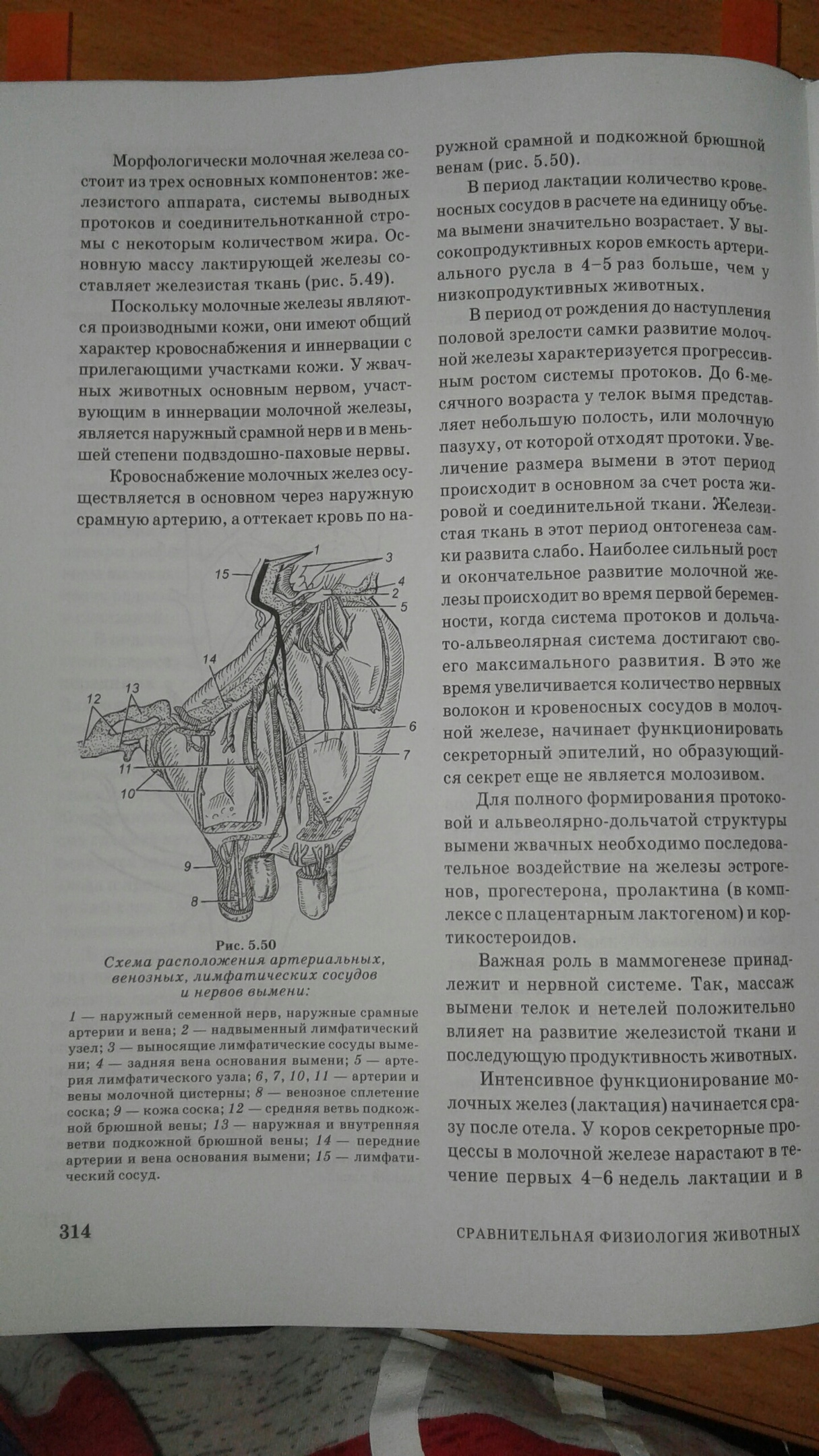
Роль печени в пищеварении.
Из всех органов печень играет ведущую роль в обмене белков, жиров, углеводов, витаминов, гормонов и других веществ. Ее основные функции:
1. Антитоксическая. В ней обезвреживаются токсические продукты, образующиеся в толстом кишечнике в результате бактериального гниения белков – индол, скатол и фенол. Они, а также экзогенные токсические вещества (алкоголь), подвергаются биотрансформации (индол в индикан, который выводится с мочой).
2. Печень участвует в углеводном обмене. В ней синтезируется и накапливается гликоген, а также активно протекают процессы гликогенолиза и неолюкогенеза. Часть глюкозы используется для образования жирных кислот и гликопротеинов.
3. В печени происходит дезаминирование аминокислот, нуклеотидов и других азотсодержащих соединений. Образующийся при этом аммиак нейтрализуется путем синтеза мочевины.
4. Печень участвует в жировом обмене. Она преобразует короткоцепочечные жирные кислоты в высшие. Образующийся в ней холестерин используется для синтеза ряда гормонов.
5. Она синтезирует ежесуточно около 15 г альбуминов, α1- и α2-глобулины, β2-глобулины плазмы.
6. Печень обеспечивает нормальное свертывание крови, α2-глобулинами является протромбин. Ас-глобулин, конвертин, антитромбины. Кроме того, ею синтезируется фибриноген и гепарин.
7. В ней инактивируются такие гормоны, как адреналин, норадреналин, серотонин, андрогены и эстрогены.
8. Она является депо витаминов А, В, D, Е, К.
9. В ней депонируется кровь, а также происходит разрушение эритроцитов с образованием из гемоглобина билирубина.
10. Экскреторная. Ею выделяются в желудочно-кишечный тракт холестерин, билирубин, мочевина, соединения тяжелых металлов.
11. В печени образуется важнейший пищеварительный сок – желчь.
Желчь вырабатывается гепатоцитами путем активного и пассивного транспорта в них воды, холестерина, билирубина, катионов. В гепатоцитах из холестерина образуются первичные желчные кислоты – холевая и дезоксихолевая. Из билирубина и глюкуроновой кислоты синтезируются водорастворимые комплексы..
Желчь вырабатывается печенью постоянно. В сутки ее образуется около 1 литра. Гепатоцитами выделяется первичная или печеночная желчь. Это жидкость золотисто-желтого цвета, щелочной реакции. Ее pH=7,4-8,6. Она состоит из 97,5% воды и 2,5% сухого остатка. Желчь выходит в двенадцатиперстную кишку. В Сухом остатке содержатся:
1. Минеральные вещества. Катионы натрия, калия, кальция, гидрокарбонат, фосфат анионы, анионы хлора.
2. Желчные кислоты – таурохолевая и гликохолевая.
3. Желчные пигменты – билирубин и его окисленная форма биливердин. Билирубин придает желчи цвет.
4. Холестерин и жирные кислоты.
5. Мочевина, мочевая кислота, креатинин.
6. Муцин.
Значение желчи:
1. Желчные кислоты эмульгируют часть жиров, превращая крупные жировые частицы в мелкодисперсные капли.
2. Она активирует ферменты кишечного и поджелудочного сока, особенно липазы.
3. В комплексе с желчными кислотами происходит всасывание длинноцепочечных жирных кислот и жирорастворимых витаминов через мембраны энтероцитов.
4. Желчь способствует ресинтезу триглицеридов в энтероцитах.
5. Инактивирует пепсины, а также нейтрализует кислый химус, поступающий их желудка. Этим обеспечивается переход от желудочного к кишечному пищеварению.
6. Стимулирует секрецию поджелудочного и кишечного соков, а также пролиферацию и слущивание энтероцитов.
7. Усиливает моторику кишечника.
8. Оказывает бактериостатическое действие на микроорганизмы кишечника и таким образом препятствует развитию гнилостных процессов в нем.
Регуляция желчеобразования и желчевыведения в основном осуществляется гуморальными механизмами, хотя некоторую роль играют и нервные. Самыми мощными стимуляторами желчеобразования в печени являются желчные кислоты, всасывающиеся в кровь из кишечника. Его также усиливает секретин, который способствует увеличению содержания в желчи гидрокарбоната натрия. Блуждающий нерв стимулирует выработку желчи, симпатические тормозят.
Важную роль в обмене белков играет печень. Часть аминокислот, поступающих с кровью, используется здесь для синтеза неспецифических белков, а часть подвергается расщеплению с образованием безазотистого остатка, используемого в дальнейшем для синтеза углеводов, а также аммиака. Последний превращается в мочевину и выводится из организма почками.
Газообмен лошади
Лошади имеют хорошо развитые легкие массой 4,5–6,5 кг и емкостью до 50 л, что позволяет им усваивать большое количество кислорода (рис. 5.13). Каждое легкое разделяется лишь сердечной вы0 резкой на краниальную и каудальную доли. Междольковая ткань развита слабо, поэтому поверхность легких гладкая. Дышат лошади только через ноздри, а поступление воздуха регулируется подвижными крыловидными хрящами. Ноздри являются уязвимым местом у лошади при таких заболеваниях, как сап, мыт. Носового зеркала у лошади нет. Верхняя губа без заметных границ переходит в спинку носа. Ноздри большие, по форме напоминают перевернутую запятую. Верхняя суженная часть ноздри служит вхо0 дом в носовой дивертикул. Мягкая боковая стенка носа и щель между началом дивертикула и носовой полостью делают возможным широкое раскрытие ноздрей при интенсивном дыхании. Число дыхательных движений (вдохов и выдохов) в состоянии покоя у лошади находится в пределах 8–16 раз в 1 мин (приложение 3). При напряженной мышечной работе на рыси и в галопе частота дыхания возрастает до 120 раз в 1 мин. Чем резвее лошадь, тем объемнее у нее легкие. При напряженной и продолжительной работе они могут увеличивать легочную вентиляцию в 10–12 раз. Так, в состоянии покоя лошадь вдыхает при каждом дыхательном цикле 5–6 л возду0 ха. При частоте дыхания 12 вдохов за 1 мин минутный объем воздуха будет составлять 60–70 л. При легкой работе минутный объем возрастает до 150–200 л, а при интенсивной работе — до 400–500 л. При изнуряющих физических нагрузках лошади способны переходить с аэробного дыхания на анаэробное. В состоянии покоя лошадь за 1 мин потребляет 1,2– 1,8 л кислорода, а после бега — 6–8 л (больше в 5,5 раз). При движении тро0 том кислорода потребляется больше в 12 раз, а при резвой рыси — в 36 раз (до 64 л). При значительном возрастании скорости движения лошади ее энергетические затраты намного превышают уровень поглощения кислорода, вследствие чего в организме животного образуется «кислородный долг». Он возникает уже при движении лошади тротом, а при резвой рыси достигает 40%, что приводит в действие буферные и резервные системы организма. Долгая напряженная работа приводит к полному истощению этих ресурсов и к гибели животного.
Кишечное пищеварения.
Содержимое желудка переходит в двенадцатиперстную кишку дробными порциями благодаря ритмичному открыванию и закрыванию пилорического сфинктера. Это осуществляется рефлекторно и носит название пилорного рефлекса. Кормовые массы, поступающие в двенадцатиперстную кишку, подвергаются воздействию со стороны сока поджелудочной железы, желчи и кишечного сока, которые имеют щелочную реакцию. В результате кислое содержимое желудка постепенно нейтрализуется и начинает формироваться энтеральный химус. Скорость прохождения химуса по кишечнику — 8,5 м за сутки. Отношение длины тонкого кишечника (20–40 м) к длине туловища у лошади составляет 12 : 1. Окончательное выделение порции корма происходит через 12–20 дней. В опытах на жеребятах с хронической фистулой поджелудочной железы и желчного протока было установлено, что секреция поджелудочного сока у лошади непрерывна. Прием корма резко усиливает отделение поджелудочного сока. За сутки поджелудочная железа лошади секретирует 7,5–8,5 л сока. Протеолитическая активность сока у лошади равняется 2,1– 2,0 мм, по Мету. Каждый вид корма способствует секреции поджелудочного сока с разной интенсивностью. Наибольшее отделение сока отмечается при кормлении лошадей пшеничными отрубями. Уже через час после поедания отрубей отделение сока увеличивается более чем в два раза. Максимальная секреция наблюдается через 3 ч. Содержание ферментов в поджелудочном соке увеличивается к четвертому часу сокоотделения. При поедании комбинации овса и сена также прослеживается увеличение секреции поджелудочного сока (максимальное увеличение отмечается через 3 ч). Одна! ко в этом случае объем выделенного сока меньше, чем при скармливании отрубей. Кроме сока поджелудочной железы, в двенадцатиперстную кишку через печеночный проток поступает желчь. Печень лошади за сутки вырабатывает 5– 6 л желчи. У лошадей отсутствует желчный пузырь, его роль в определенной мере выполняет так называемая желчная цистерна, т. е. расширение общего печеночного протока. Из-за отсутствия желчно! го пузыря, слизистая оболочка которого у других животных вырабатывает слизь, примешивающуюся к желчи, желчь у лошади имеет более жидкую консистенцию. Собственно кишечный сок — это секрет бруннеровых и либеркюновых желез. Физико-химические свойства кишечного сока у лошади отражают следующие показатели: рН — 8,16–8,68; вязкость — 11,16–17,66; сухое вещество — 1,99– 5,48%; удельный вес — 1,004–1,014; азот общий — 1,92–6,06 мг%; азот аммонийный — 0,358–0,661 мг%. После прохождения тонкого отдела кишечника химус поступает в толстый отдел кишечника. Пищеварение в этом отделе ЖКТ у лошади в основном происходит за счет ферментов, поступающих сюда вместе с химусом из вышележащих отделов. В толстом отделе кишечника к ферментам, как основным факторам переваривания корма, присоединяется бактериальная флора. Бактерии в ЖКТ поступают с кормом. При прохождении по пищеварительному тракту корм смачивается пищеварительными соками, имеющими бактерицидное действие (слюна, желудочный и поджелудочный сок, желчь). Благодаря быстрому продвижению химуса и бактерицидности соков ни в желудке, ни в тонком кишечнике бактерии не имеют благо! приятных условий для своего развития. И только в толстом отделе вследствие снижения перистальтики содержимое задерживается на более продолжительное время, что создает благоприятные условия для размножения бактерий. Под действием микроорганизмов здесь происходят процессы гидролиза и гниения белков, сбраживания углеводов и окисления жиров, а также образования ряда газообразных продуктов, таких как Н2, Н2S, CH4, CO2 и NH3. Появляющиеся при распаде белков ядовитые продукты — фенол, ин! дол, скатол и другие — нейтрализуются в печени и выводятся из организма ло! шади с мочой и калом. Особое значение в пищеварении лошади имеет слепая кишка. Ее объем составляет 32–37 л, что примерно вдвое больше желудка. Кормовые массы, богатые ферментами, поступают сюда из тон! кого кишечника. Ферменты кишечного сока продолжают свое действие на субстраты в слепой кишке. Наряду с этим в слепой кишке происходит микробный процесс сбраживания клетчатки, имеющий особое значение для лошади как травоядного животного. Образующиеся в результате брожения клетчатки летучие жирные кислоты (ЛЖК) поступают в кровь и используются для синтеза и белка. Соотношение ЛЖК в содержимом слепой кишки у лошади составляет: уксусная кислота — 73%, пропионовая кислота — 20%, маслянная и другие кислоты — 7%. Характер переваривания корма в слепой кишке у лошади напоминает изменения корма в рубце у жвачных. Только образующиеся кислоты нейтрализуются щелочной слизью, вырабатываемой железами слизистой оболочки слепой кишки. Щелочная слизь нормализует бродильные процессы в кишке. Из слепой кишки содержимое транспортируется в большую ободочную кишку. В большой ободочной кишке пищеварительные процессы напоминают таковые слепой кишки. Однако они протекают с меньшей интенсивностью. В малой ободочной кишке реакция содержимого становится кислой, процессы гидролиза прекращаются и пищевые массы приобретают фекальный вид и запах, типичные для лошади. Содержимое в слепой и ободочной кишке лошади задерживается до 72 ч. Это благоприятствует процессам расщепления основных нутриентов рациона. Здесь переваривается до 50% всей клетчатки, до 40% белка, до 25% крахмала и сахаров. Для толстого отдела кишечника лошади характерна своеобразная моторика. В дистальном отделе кишечника появляются глубокие кольцевые сокращения стенки кишечника, которые распространяются в каудальном направлении. Кольцевые сокращения кишки уплотняют химус и формируют каловые массы. Это обеспечивает своеобразную форму кала у лошадей. Они же обеспечивают и дефекацию, т. е. удаление кала из прямой кишки. У лошади за сутки выделяется до 20 кг кала. Формирование кала происходит в нижних отделах толстого кишечника, где всасывается вода. Кал представляет собой остатки непереваренного корма с примесью слизи, желчи. От 30 до 50% массы свежего кала лошади приходится на микробы — микрофлору и простейших
Системы анализаторов.
Зрительный анализатор
Из сухопутных животных у лошади самые большие глаза (диаметр глазного яблока 51 мм). Они расположены по бокам головы, чем обеспечивается довольно значительное поле зрения (на 180°). У лошади зрачок поперечно-овальной формы, на верхнем крае его имеется 2 – 4 плотных образования, укрепленных на ножках («виноградные зерна»). Угол между осями глазниц составляет 115°, а между зрительными осями 137°, т.е. у лошади бинокулярное объемное зрение, дающее возможность видеть не только предметы, но и различать их форму, величину и соотношения друг к другу. Глаз у лошади астигматический, т.к. роговица имеет не шаровидную, а эллипсоидную форму, т.е. в различных направлениях она имен неодинаковый радиус кривизны. По краям роговицы преломление света иное, чем в центре, поэтому лошадь может видеть даже очень мелкие предметы. В связи с анатомическими особенностями строения глаза оценка пространственных ситуаций у лошади затруднена. Для получения точной картины на сетчатке лошадь должна выпрямить или склонить голову. Лошадь хорошо видит ночью, однако адаптационные свойства глаза к свету и темноте значительно ниже, чем у человека и ночных животных. Лошади обладают очень большой остротой зрения и способны к довольно тонкой дифференцировке световых раздражителей. Они хорошо отличают источники света яркостью 1808 от яркости в 1903 кд/м2. Электрофизиологические исследования показали, что лошади обладают цветовым зрением. Они способны различать почти все цвета. Имеется еще интересная особенность. Лошади воспринимают как живых чучело, скульптуру, портрет животных или человека.
Слуховой анализатор
Установлено что слуховой анализатор лошади высоко развит и способен воспринимать, анализировать и синтезировать разнообразные звуковые раздражители, и в этом отношении лошади почти не уступают собакам. Острота слуха лошади значительно превосходит остроту слуха человека. Ухо человека воспринимает от 16000 до 20000 колебаний (Гц с секунду), а ухо лошади до 40000 Гц/с., т.е. ухо лошади улавливает шорохи, не воспринимаемые человеческим ухом. Этому способствует значительная подвижность ушных раковин, повышающая восприятие звука и воронкообразное строение их, улучшающее улавливание звуковых волн и усиливающее звуковые раздражения. Лошади способны воспринимать звуки в диапазоне от 200 до 21000 Гц. Однако предполагают, что верхняя граница слуха лошади лежит где-то около 30 – 40000 Гц. Причем острота слуха лошади на разную частоту звука неодинакова: она большая на частоту от 1 до 6000 Гц и наименьшая на все остальные тоны. Лошади обладают высокой способностью дифференцировать звуковые раздражители, что зависит от типологических особенностей их ВНД. Так, лошади сильного уравновешенного подвижного типа способны более тонко дифференцировать, чем лошади безудержного и слабого типов.
Обонятельный анализатор
Лошадь относится к макросматикам, у нее обоняние развито лучше, чем у других видов домашних животных. Значительная часть переднего мозга у них связана с функцией обоняния. Лошадь ориентируется по следу животных так же хорошо, как и собака. Ее устрашает запах крупных хищников, волнует запах крови, настораживает табачный дым. Она безошибочно находит пути к дому и водным источникам на расстоянии до 10 км. Лошадь определяет по запаху малейшие примеси в воде. Лошади не выносят резких и дурных запахов, в том числе запахов духов и водки. Они отказываются поедать корм, пить воду, если от кормушек, ведер пахнет дезсредствами. Редко ими поедаются прелый, заплесневевший овес, сено. Лошади всех возрастов не поедают белену, дурман, репейник, лопух, конский щавель и другие ядовитые травы в любой стадии развития. На остроту обоняния у лошадей влияют температура и влажность воздуха, атмосферное давление и др. С появлением росы, тумана лошади перестают пастись на пастбище, ибо при этом у них снижается обоняние.
Вкусовой анализатор
Вкусовая рецепция у лошади тесно связана с обонятельной и развита лучше, чем у других животных. Она очень разборчива в кормах, не ест многих трав, хорошо отбирает в кормушке зерна овса от зерен куколя. Лошади любят осину, кора которой отличается сильной горечью, менее приятно для них кислое. Вкусовое чувство у новорожденных жеребят пробуждается раньше всех остальных ощущений.
Двигательный анализатор
Двигательный анализатор развит очень хорошо и тесно связан с другими анализаторами. Так, движения у лошади очень легко связываются с ритмичным звуковым раздражителем, т.к. такие движения, как ходьба, бег являются ритмичными. Зрительный анализатор обеспечивает пространственную динамику мышечной активности. Вестибулярный анализатор взаимодействует с двигательным при формировании и осуществлении двигательных актов при изменении положения тела в пространстве.
КОЖНЫЙ АНАЛИЗАТОР
Развит хорошо. Особенно отчетливо выражена болевая и тактильная чувствительность у лошади в области головы (верхняя губа, нос, веки, кончик языка) и копытного венчика, т.к. в этих участках рецепторы расположены более концентрированно. Лошадь хорошо чувствует дорогу копытом. На губах, вокруг глаз, ноздрей, на ушных раковинах расположены специальные длинные и упругие осязательные волоски – вибрисы. Тактильная рецепция хорошо развита на всей поверхности кожи. Лошади могут тонко дифференцировать положительные тактильные раздражители (касалки) от отрицательных при нанесении их на кожу на расстоянии 3 см друг от друга. Они способны довольно точно определять место локализации раздражений, например, полжанье насекомых по коже и т.п. У лошади условные рефлексы на кожно-механические раздражители вырабатываются быстрее, чем на любые другие.
Состав крови свиней
Биохимические показатели крови свиней, отражая особенности промежуточного обмена веществ, находятся под контролем нервной и эндокринной систем. Наиболее важными показателями, характеризующими напряженность обмена веществ и физиологическое состояние свиней, являются белковый состав сыворотки крови и уровень содержащейся в ней глюкозы. Состав крови зависит от физиологического состояния организма, возраста, пола, условий кормления свиней и других факторов. В цельной крови свиней содержится 20% сухого вещества, в плазме— 9%. Плотность крови свиней — 1,046 (1,039-1,054), плазмы — 1,022- 1,033. Плотность крови-поросят такая же как и у взрослых животных. Она может увеличиваться при сгущении крови и уменьшаться при анемиях.В частности железодефицитная анемия возникает по разным причинам. Среди них затрудненное всасывание железа в желудочно-кишечном тракте. Одной из причин развития железодефицитной анемии служит повышенное расходование железа при хронических кровопотерях, вызванных заболеваниями, повышенной лактацией, интенсивной мышечной работой. У свиней плазма крови представляет собой прозрачную жидкость с желтоватым оттенком. Она состоит на 91% из воды и на 9% из сухого вещества. Сухое вещество плазмы в основном представлено белками и неорганическими солями. Все белки плазмы связывают катионы крови, переводя их в недиффундирующую форму. У свиноматок в плазме крови содержится в среднем 7,24-8,56% белка. Белки плазмы крови можно разделить на несколько фракций — альбумины (45%), а- (17%), в- (18%) и у-глобулины (20%). Сдвиги в обмене веществ отражаются на белковом составе плазмы крови. У новорожденных поросят и молодняка старшего возраста в плазме содержится меньше общего белка, у-глобулинов и всех аминокислот, а с возрастом их количество увеличивается. Кроме белка в плазме крови имеются небелковые азотсодержащие соединения — мочевина, аммиак, креатинин. Совокупность этих веществ объединяют под названием «остаточный азот», которыйобнаруживается в крови свинейв значительном количестве. Увеличение остаточного азота в крови у свиней происходит при инфекциях, дистрофии печени, почечной недостаточности, нефритах и переломах костей, а снижение его уровня происходит при печеночной недостаточности, дистрофии, циррозе печени, а у свиноматок — во время супоросности. Количество общего белка, белковых фракций и свободных аминокислот в плазме крови у свиней разных пород неодинаково. В плазме крови свиней имеются безазотистые органические вещества — углеводы, жиры и продукты их распада. Углеводы крови – глюкоза. Количество глюкозы в крови у свиней 58-140 мг на 100 мл крови. С возрастом содержание глюкозы в крови свиней падает, что отражает снижение уровня обменных процессов в организме животных. У более мелких свинок уровень глюкозы выше. После опороса в крови свиноматок содержание глюкозы повышается на 1,5- 6,0 мг%, т.к. обмен энергии у свиней в подсосный период возрастает. Новорожденные поросята имеют ограниченные_запасы легкодоступной энергии. У новорожденных поросят в крови содержится 10-100 мг% глюкозы, уровень которой ко 2-му дню жизни снижается. Голодание новорожденных поросят в течение первых 2 дней практически исчерпывает все запасы глюкозы в крови и является причиной падежа.
Регуляция лактации.



Насекомые
Биологические особенности насекомых и их роль в природе
1. Многослойная хитинизированная кутикула с наружным слоем, содержащим воскоподобные и жировые вещества, защищающая тело от потери влаги, механических повреждений, воздействия ультрафиолетовых лучей.
2. Развитие с метаморфозами. Выделяют следующие стадии: яйцо — личинка — куколка — имаго (взрослое половозрелое насекомое). Есть виды животных осуществляющих неполный метаморфоз, в их развитии нет стадии куколки. Личинка этих насекомых представляет собой миниатюрную копию взрослого насекомого, постепенно от линьки к линьке увеличивающегося в размерах. Неполный метаморфоз свойственен таким насекомым, как тараканы, сверчки, клопы и кузнечики. Насекомые, с полным метаморфозом, находясь в стадии куколки, претерпевают более глубокую перестройку органов. Наиболее сложен этот процесс у мух, у которых после окукливания большая часть внутренних органов личинки приобретает кашицеобразную консистенцию.
3. Конечности насекомых - система рычагов, позволяющая совершать разнообразные и точные движения. Большая часть видов насекомых приспособлена к полету. Бескрылость связана с утратой крыльев и свойственна преимущественно паразитическим видам.
4.Способность к полету, позволяющая насекомым быстро заселять новые территории, преодолевать водные пространства и другие преграды; большая подвижность, обеспеченная развитой поперечнополосатой мускулатурой, членистыми конечностями.
5. Фасеточные глаза — состоят из большого числа простых глазков, количество которых велико у активных хищников и мало у обитателей подземелий. К примеру, глаз стрекозы состоит из 28000 фасеток, а муравья — всего из 8. Помимо фасеточных глаз у насекомых есть и примитивные глазки, расположенные в различных частях тела и воспринимающие только интенсивность освещения.
5. Трахейная дыхательная система — сложная сеть трубочек, разносящих кислород ко всем органам и тканям. Трахеи позволяют осуществлять интенсивный газообмен и поддерживать при необходимости (во время полета) высокий уровень процессов жизнедеятельности.
6. Способность к мимикрии. Окраска многих насекомых изменяется в зависимости от цветового окружения, а представители некоторых видов и по форме напоминают веточки дерева.
7. Анабиоз и диапауза. Анабиоз — это оцепенение, вызванное замедлением всех жизненных функций из-за снижения температуры. Существует критическая температура (-12 градусов), охладившись ниже ее, организм насекомого может погибнуть. Но существует защитный механизм, срабатывающий при опускании температуры ниже -12 градусов и затвердевании гемолимфы организма насекомого, он заключается в резком высвобождении резервной энергии, повышающей температуру тела насекомого до 0 градусов. Этот защитный механизм эффективен при непродолжительном однократном снижении температуры. Если же температура снова опускается ниже критической, организм гибнет. При нагревании воздуха выше +40 градусов насекомые впадают в тепловое оцепенение — диапаузу. Температура, превышающая +52 градуса губительна для насекомых с их несовершенными механизмами теплоотдачи.
8.Разнообразие ротовых аппаратов, позволяющее использовать различный кормовой материал, что уменьшает межвидовую конкуренцию и способствует поддержанию высокой численности насекомых.
9.Малые размеры насекомых, обеспечивающие выживание и способствующие созданию необходимых условий для существования даже в очень незначительных по размеру пространствах (небольшие обрастания на скалах, трещины в коре деревьев, почве и др.).
10.Разнообразие способов размножения — кроме обоеполого, размножение партеногенетическое (у тлей), на стадии личинок (у отдельных видов двукрылых, жуков и клопов). Некоторым паразитическим перепончатокрылым свойственна полиэмбриония (бесполое размножение на стадии делящейся зиготы), позволяющая им резко увеличивать численность потомков.
11.Высокая плодовитость и способность к массовому размножению : среднее число откладываемых яиц составляет 200— 300. Некоторые насекомые откладывают небольшое число яиц, но дают несколько поколений (до 10 и более) за вегетационный сезон. Эта способность вызывает массовое появление многих насекомых (хрущи, мухи, комары, саранча и др.).
12.Разнообразие типов постэмбрионального развития : неполный, полный метаморфоз и другие типы превращения. В фазе личинки происходит рост и развитие особи, в фазе взрослой особи — размножение и расселение. Способность переживать неблагоприятные условия в состоянии диапаузы — временного физиологического покоя.
13.Смена среды обитания на разных стадиях онтогенеза: личинки обитают в водной среде, взрослые — в наземно-воздушной (например, стрекозы, комары и др.), что снижает внутривидовую конкуренцию за пищу, пространство для жизни и способствует лучшему выживанию насекомых.
14. Хорошо развитая нервная система, разнообразные и совершенные органы чувств, сложные врожденные формы индивидуального и общественного поведения — инстинкты. Особенности строения и процессов жизнедеятельности. Тело насекомых разделено на голову, грудь и брюшко. На голове находится пара членистых усиков, пара верхних и две пары нижних челюстей. Кроме того, они имеют пару сложных фасеточных глаз, а многие — и простые глазки. Строение ротового аппарата разнообразно и соответствует характеру питания. Исходным считается грызущий ротовой аппарат, от которого в результате пищевой специализации произошли остальные типы: колюще-сосущий (у комаров, клопов, тлей и др.), сосущий (у чешуекрылых), лакающий (у пчел, шмелей), лижущий (у мух). Грызущим ротовым аппаратом обладают, в частности, насекомые отрядов жестко - и прямокрылых, а также многие личинки.
Роль насекомых: Насекомые участвуют в почвообразовательных процессах, уничтожают отмершие части растений и вносят их в почву, обогащая перегноем. Уничтожение трупов и экскрементов имеет большое санитарное значение. Насекомые опыляют цветковые растения. Главную роль среди опылителей играют перепончатокрылые, двукрылые и чешуекрылые. Насекомые дают ценную пищевую и техническую продукцию, например пчёлы – мёд, воск и другие продукты. Из коконов тутового шелкопряда вырабатывают натуральный шёлк. Многие насекомые служат пищей позвоночных животных. Их поедают птицы, млекопитающие, рыбы, амфибии, рептилии. Энтомофаги сдерживают увеличение численности вредителей сельского и лесного хозяйства. Эту функцию выполняют хищные жуки, перепончатокрылые и двукрылые, наездники и яйцееды. Насекомые имеют большое эстетическое значение. Многие виды бабочек и жуков, стрекоз и перепончатокрылых являются украшением природы. Насекомые играют большую роль в природе и жизни человека. Основная пища большинства их – это разные части растений (от листьев, плодов, семян, пыльцы и нектара цветов до древесины). Поэтому насекомые имеют особенно большое значение во всех наземных растительно – животных группировках как важнейшие потребители растений. Из-за этого они нередко оказываются очень опасными вредителями сельского хозяйства.
Дата: 2019-02-19, просмотров: 877.