Brendemoen [41], Comoens и соавт. [58] и другие авторы [199, 224] обнару-жили, что антигены Le a и Le b на эритроцитах беременных выражены слабее, чем до беременности. Некоторые женщины, типированные как Le(a −b + ) или Le(a +b −), во время беременности приобретали фенотип Le(a −b −), т. е. полно-стью утрачивали антигены Lewis. В то же время способность слюны этих жен-щин нейтрализовать сыворотки анти-Le a и анти-Le b не нарушалась (Hammar и соавт. [104] , Taylor и соавт. [224]).
На ослабление групповых антигенов АВО при беременности указывали Schachter и соавт. (цит. по Issitt, Anstee [115]), объясняя это тем, что продукция А-генспецифической N-ацетилгалактозаминилтрансферазы у беременных за-метно снижается, вследствие чего уменьшается синтез группоспецифических олигосахаридов. Подобное объяснение экстраполировано на редукцию у бере-менных антигенов Lewis. Однако наблюдения Hammer и соавт. [104] показа-ли, что это не совсем так. У беременных уровень олигосахаридов Lewis в плаз-ме почти такой же, как у небеременных, но относительная концентрация липо-протеинов по сравнению с общей клеточной массой эритроцитов существен-но выше. Вновь синтезируемые олигосахариды Lewis, по-видимому, с боль-шей скоростью связываются с липопротеинами плазмы, чем со сфинголипида-ми стромы эритроцитов. При таких условиях на эритроцитах адсорбируется су-щественно меньше Lewis-олигосахаридов, чем вне беременности.
Маловероятно, что беременные могут вырабатывать антитела анти-Le a и анти-Le b в тот период, когда антигены Lewis на их эритроцитах отсутству-ют. Тем не менее остается фактом, что частота обнаружения антител анти-Le a
80. анти-Le b у беременных выше, чем у других лиц (Kissmeyer-Nielsen [132]). В этом проявляется еще одна особенность системы Lewis.
Issitt и Anstee [115] привели интересный случай: у женщины на 7-м мес. бе-ременности обнаруживали сильные антитела анти-Le b, но когда через 3 мес. по-сле рождения ребенка у нее была вновь взята кровь с целью получения тестово-го реактива анти-Le b, антител в ее сыворотке не оказалось, а эритроциты имели фенотип Le(a −b + ).
567
Комментируя этот случай, Issit указывает, что частота обнаружения антител против антигенов Lewis могла бы быть неизмеримо выше при условии целена-правленного скрининга этих антител. Однако скрининг антител Lewis в родов-спомогательных учреждениях и учреждениях службы крови, как правило, не проводят из-за малой их клинической значимости.
Внезапное исчезновение антител Lewis, по-видимому, возможно не только у беременных женщин. Мы наблюдали исчезновение антител анти-Le a у 22-лет-него мужчины через год с момента их обнаружения [5].
Фенотип Le (a +b + )
Среди европеоидов этот фенотип встречается редко, но чаще всего у пред-ставителей монголоидных рас (японцев [146, 218], тайцев [56]), а также у не-гроидов и австралоидов (аборигенов Австралии [36, 232], Полинезии [108]).
Broadberry и Lin-Chu [44] нашли фенотип Le(а +b + ) у 22 −25 % китайцев жи-телей Тайваня; Henry и соавт. [108] − у 10 −40 % полинезийцев.
106. соответствии с существующими представлениями о синтезе олигосаха-ридов Lewis фенотип Le(a +b + ) возникает, если Le-генспецифическая транс-фераза сверхактивна и добавляет L-фукозу к субтерминальному N-ацетил-D-глюкозамину прежде, чем Se-генспецифическая трансфераза присоединит L-фукозу к терминальной D-галактозе. В результате антиген Le a не может быть конвертирован в Le b и остается в плазме в виде олигосахарида Le a. Наряду с этим синтезируется и олигосахарид Le b. Оба олигосахарида (Le a и Le b) при-сутствующие в плазме, адсорбируются на эритроцитах, придавая им фенотип
Le(a +b + ).
Henry и соавт. [107, 108] пришли к выводу, что фенотип Le(a +b + ) появляет-ся в результате действия особого гена Se w (аллеля Se), являющегося суперактив-ной формой гена Le.
Наличие гена Se w позволило Cowles и соавт. [60] и другим авторам [44, 108] объяснить повышенную частоту фенотипа Le(a +b + ) среди жителей Тайваня.
Обнаружены две мутации гена Se: одна влияет на активность гена Se у евро-пейцев, обе вместе изменяют активность гена Se у полинезийцев несекреторов.
Ген S w пока детально не изучен. Не исключено, что фенотип Le(a +b + ), об-условливаемый геном Se w, отчасти может быть связан с тем, что типирование монголоидов проводят сыворотками европеоидов, которые, как нам представ-ляется, могут неодинаково реагировать с эритроцитами представителей разных рас. Однако это всего лишь наше предположение.
Эритроциты, адсорбирующие Le b из плазмы и являющиеся Le(a −b + ), часто несут некоторое количество вещества Le a. Обычные поликлональные сыворот-ки анти-Le a и анти-Le b типируют такие клетки как Le(a −b + ), однако сильные сыворотки анти-Le a, особенно моноклональные, способны выявить антиген Le a.
Некоторые авторы [115] полагают, что при исследовании различных по-пуляций, в том числе монголоидных, с помощью активных моноклональных
568
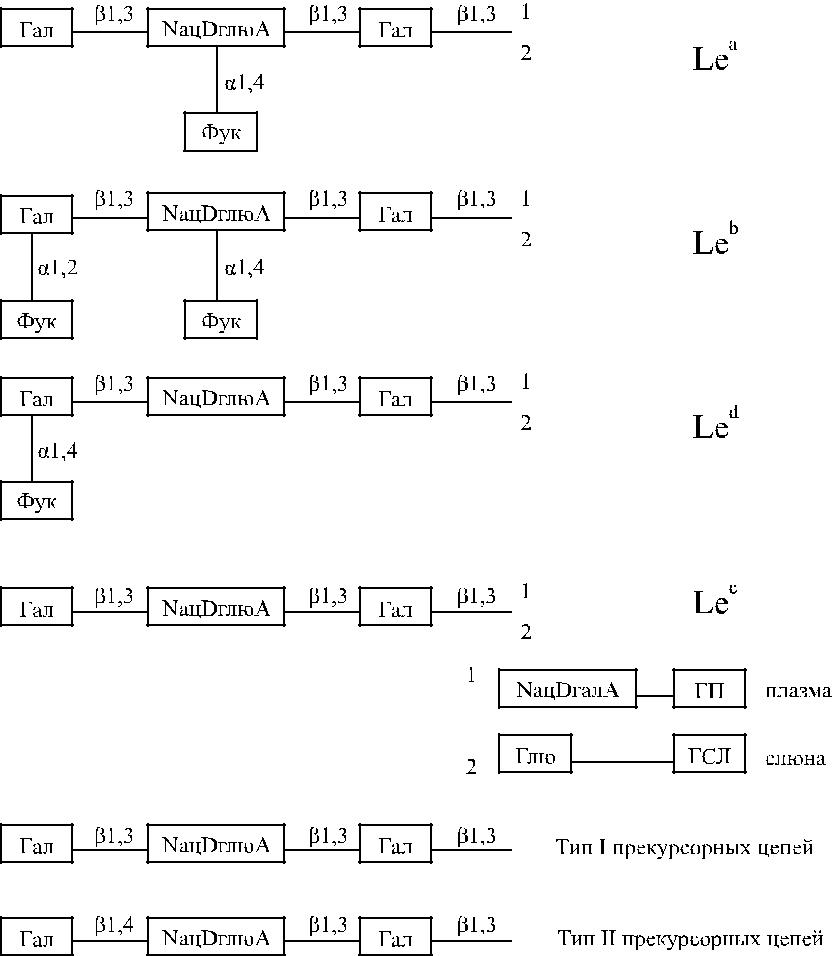
Рис. 9.1. Химическая структура антигенов Le а, Le b, Le d и Le c.
NацDгалA − N-ацетил-D-галактозамин
NацDглюA − N-ацетил-D-глюкозамин
Гал − D-галактоза
Глю – глюкоза
ГСЛ − гликосфинголипиды ГП − гликопротеины Фук − L-фукоза
реагентов анти-Le a, следовые количества Le a, часто присутствующие, но не вы-являемые поликлональными сыворотками анти-Le a, могли создавать видимость фенотипа Le(a +b + ). Вместе с тем нельзя полностью исключить возможность существования у монголоидов гена Se w, обусловливающего более высокую ча-стоту фенотипа Le(a +b + ) по сравнению с европеоидными популяциями.
Sturgeon и Arcilla [219], обследуя семьи, где имелись родители и дети с фенотипом Le(a +b + ), констатировали, что у лиц Le(a +b + ) реакция с Le а вы-ражена сильнее, чем с Le b. В слюне обнаруживали сильные субстанции Le x
88. Н. У 13 контрольных доноров Le(a +b −) в слюне отсутствовали субстан-ции Le b и АВН. По содержанию веществ Le а и Le b в слюне лица с фенотипом Le(a +b + ) занимали промежуточное положение между лицами Le(a −b −), имеющими низкий титр групповых субстанций, и Le(a −b + ) с высоким ти-тром групповых веществ.
569
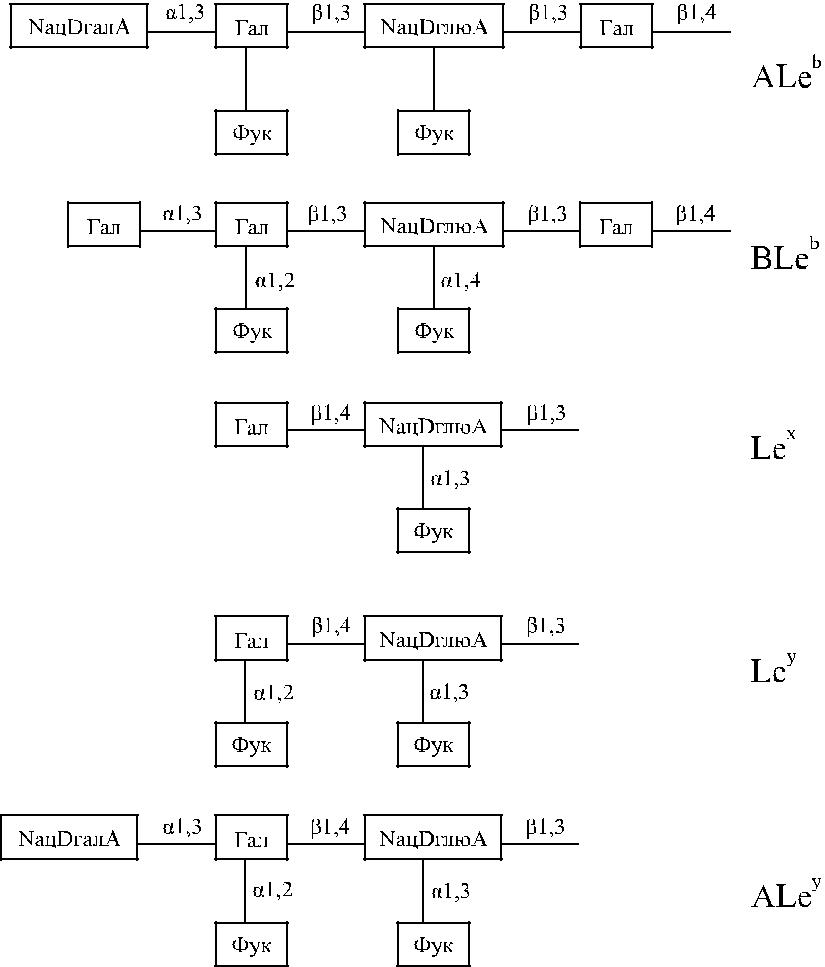
Рис. 9.2. Химическая структура антигенов ALeb, BLeb, Lex, Ley и ALey. NацDгалA – N-ацетил-D-галактозамин ГСЛ – гликосфинголипиды
NацDглюA – N-ацетил-D-глюкозамин ГП – гликопротеины
Гал – D-галактоза Фук – L-фукоза
Глю – глюкоза НейК – нейраминовая кислота
Дата: 2019-02-24, просмотров: 467.