(из статьи К. Ландштейнера [135])
| RBC | Dr.St. | Dr.Ples. | Dr.Sturl. | Dr.Erdh. | Mr.Zar. | Mr.Land. |
| Serum | ||||||
| Dr.St. | − | + | + | + | + | − |
| Dr.Ples. | − | − | + | + | − | − |
| Dr.Sturl. | − | + | − | − | + | − |
| Dr.Erdh. | − | + | − | − | + | − |
| Mr.Zar. | − | − | + | + | − | − |
| Mr.Land. | − | + | + | + | + | − |
Позднее была открыта группа АВ, сыворотки крови таких лиц не содержали изогемагглютининов, в то время как их эритроциты несли антигены А и В одно-временно (Decastello, Sturli [90]).
Dungern и Hirszfeld [94], проведя посемейные исследования, установили пе-редачу указанных признаков по наследству. В 1924 г. Bernstein [76] предложил генетическую модель, объяснявшую передачу аллелей АВО по наследству и их фенотипические проявления.
Система АВО представлена двумя антигенами: А и В. В качестве самостоя-тельных специфичностей в ней также выделяют антигены А1 и А,В. Отсутствие на эритроцитах указанных антигенов обозначают цифрой 0 или буквой О. Антитела анти-А и анти-В, обозначаемые греческими буквами α и β, имеют естественное происхождение.
64
Различают 4 группы крови, образуемые сочетаниями антигенов А, В и изогемаг-глютининов α и β. На эритроцитах лиц первой группы, О(I), антигены А и В отсут-ствуют, в плазме крови присутствуют антитела α и β. У людей со второй группой крови, A(II), на эритроцитах имеется антиген А, в плазме присутствуют антитела β. Индивиды с третьей группой, B(III), имеют на эритроцитах антиген В, в плазме при-сутствуют антитела α. У лиц с четвертой группой, AB(IV), эритроциты несут антиге-ны А и В, изогемагглютинины α и β в сыворотке крови отсутствуют (табл. 3.2).
| Таблица 3.2 | ||||||||||||
Фенотипы АВО, определяемые при рутинном типировании
Сыворотка
Реакция сывороток
Эритро-
Реакция эритроцитов с
Фено-
с эритроцитами
изогемагглютининами
Генотип
крови
циты
тип
О
А
В
αβ
β
α
Примечание. + агглютинация эритроцитов, – агглютинация отсутствует.
Агглютинационные свойства эритроцитов А(II) варьируют в широких пре-делах, в связи с чем антиген А подразделяют на А1 – сильный, высокоавидный (частота 80 %) и А2 – слабый, низкоавидный (частота 20 %) (Е.И. Дрямина [26], П.Н. Косяков [35], П. Проданов [55], Economidou и соавт. [95], Landsteiner и со-авт. [133, 134]). Таким образом, фенотипы А и АВ по интенсивности реакций
с антителами анти-А подразделяются на А1 и А2, а также А1В и А2В соответ-ственно. Антигены А и В тесно связаны с антигеном Н, который идентифици-руется специфическими антителами. Он представляет самостоятельную груп-повую систему, поскольку генные локусы АВО и Н независимы один от друго-
го (Daniels [87], Issitt, Anstee [122]). Антитела анти-А и анти-В (изогемагглюти-
нины α и β) имеют естественное происхождение и относятся к классу IgM. Они вызывают агглютинацию эритроцитов в солевой и коллоидной среде, более ак-тивны при температуре 4–20 оС, однако проявляют свое действие и при 37 оС (В.А. Аграненко, Н.Н. Скачилова [3], Н.И. Васильев и др. [7], П.Н. Косяков [35], Daniels [87], Issitt и Anstee [122]). В сыворотках крови многих людей, помимо IgM, присутствуют антитела IgG той же специфичности. Эти антитела лучше проявляют агглютинирующую активность в коллоидной среде и более активны при 37 оС (Issitt и Anstee [122], Mollison и соавт. [159]). Помимо реакции агглю-
тинации, антитела IgG могут быть выявлены в непрямом антиглобулиновом ме-
тоде (М.А. Умнова [12], Issitt и Anstee [122]).
Наряду с анти-А- и анти-В-антителами в системе АВО выделяют антите-ла анти-А,В и анти-А1, отличающиеся от анти-А и анти-В и открывающие ан-тигены, обозначаемые в соответствии с номенклатурой ISBT как АВО3 и АВО4 (Daniels [87]). Сыворотки крови лиц группы О содержат обе указанные
65
разновидности антител в дополнение к анти-А и анти-В (Н.М. Михайлова и др. [45, 47]). Антитела анти-А1 содержатся одновременно с анти-А в сыворот-ках крови лиц группы В. Антитела анти-Н содержатся в сыворотках крови лиц, имеющих редкую группу крови Оh (тип Бомбей), но иногда встречаются в сыво-ротках крови лиц, имеющих группу крови А и АВ. Они относятся к холодовым агглютининам, имеют низкий титр, при 37 оС не активны (Issitt и Anstee [122]).
Наследование
В 1924 г. Bernstein [76] предложил генетическую модель системы АВО. В со-ответствии с ней существуют 3 аллельных гена: О, А и В (в антропологии их обо-значают как r, p и q соответственно). Ген А контролирует синтез антигена А, ген
В – антигена В, ген О – молчащий – не кодирует синтеза какого-либо антигена, определяемого серологически. Лица группы О(I) всегда гомозиготны по аллелю
О (генотип О / О). Люди с группой АВ(IV) гетерозиготны по генам А и В (генотип А / В). Индивиды, имеющие группу А и В, могут быть гомо- (А / А и В / В) и гетеро-зиготными (А / О и В / О). Серологическими методами дифференцировать гомо- и гетерозиготность не удается. Тем не менее с определенной степенью вероятности можно сделать заключение о том, к какой из этих категорий относится данный индивид. Эритроциты гетерозигот содержат меньше антигенного материала, чем гомозиготы, что проявляется в меньшей агглютинабельности эритроцитов. Кроме того, эритроциты гетерозигот сильнее агглютинируются сыворотками анти-Н.
Групповую принадлежность членов семьи учитывают в судебной медицине при проведении экспертизы при спорном отцовстве или замене детей (Прокоп и Гёллер [56], А.К. Туманов и В.В. Томилин [60], А.К. Туманов [59], Yoshida [241]). В частности у родителей AA × AA не может быть детей с группой кро-ви B(III) или O(I). Группа O(I) исключена также, если один из родителей имеет группу крови AB(IV) (табл. 3.3).
Цис-АВ
В 1964 г. Seyfried и соавт. [194] описали польскую семью, в которой родите-ли А2В × O имели детей А2В и О (рис. 3.1), что противоречило правилу насле-дования групп крови: дети в данной семье должны были иметь группу крови А или В. Полученные данные позволили предположить, что, как редкое исключе-ние, гены А и В могут располагаться на одной хромосоме, в позиции цис, и пе-редаваться в таком виде потомству.
При обследовании 1 млн японских доноров группа АВ выявлена у 112 710, из них 14 индивидов унаследовали цис-АB. Подобный тип наследования груп-пы крови был обнаружен у израильтян, корейцев и представителей других на-
циональностей (Bennett и соавт. [74], Cho и соавт. [84], Fukumori и соавт. [107], Hummel и соавт. [121], Misfud и соавт. [154], Pacuzska и соавт. [181], Tzeng и со-авт. [212], Yoshida и соавт. [243, 244]).
66
Таблица 3.3
Варианты наследования группы крови
| Фенотип | Генотип | Возможный фенотип (генотип) детей | |||||||
| родителей | родителей | ||||||||
| A × A | AA × AA | A (AA) | |||||||
| AA × AO | A (AA или АО) | ||||||||
| AO × AO | А (АА или АО) или О (ОО) | ||||||||
| B × B | BB × BB | В (ВВ) | |||||||
| BB × BO | В (ВВ или ВО) | ||||||||
| BO × BO | В (ВВ или ВО) или О (ОО) | ||||||||
| AB × AB | AB × AB | А (АА) или В (ВВ) или АВ (АВ) | |||||||
| O × O | OO × OO | О (ОО) | |||||||
| AA × BB | АВ (АВ) | ||||||||
| A × B | AO × BB | АВ (АВ) или В (ВО) | |||||||
| AA × BO | АВ (АВ) или А (АО) | ||||||||
| AO × BO | АВ (АВ) или А (АО) или В(ВО) или О (ОО) | ||||||||
| A × O | AA × OO | А (АО) | |||||||
| AO × OO | А (АО) или О (ОО) | ||||||||
| A × AB | AA × AB | АВ (АВ) или А (АА) | |||||||
| AO × AB | АВ (АВ) или А (АО) или В (ВО) | ||||||||
| B × O | BB × OO | В (ВО) | |||||||
| BO × OO | В (ВО) или О (ОО) | ||||||||
| B × AB | BB × AB | В (ВВ) или АВ (АВ) | |||||||
| BO × AB | АВ (АВ) или А (АО) или В (ВВ или ВО) | ||||||||
| AB × O | AB × OO | А (АО) или В (ВО) | |||||||
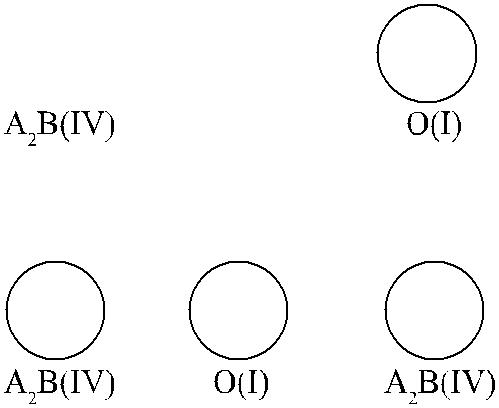
Рис. 3.1. Наследование аллеля цис-АВ.
Серологические свойства эритроцитов цис-АB несколько отличаются от обычных эритроцитов АВ (транс-АВ). Антиген А на них выражен слабее, од-нако его экспрессия выше, чем на эритроцитах А2В. Антиген В выражен слабо и представлен, как полагают Pacuzska и соавт. [181], парциальной формой, на-поминающей вариант В 3 или BW. В отдельных случаях экспрессия антигена В у лиц цис-АB нормальная (Issitt и Anstee [122]).
67
При адсорбции иммунных анти-В-антител эритроцитами цис-АB полностью удаляется фракция, реагирующая с указанными клетками. Антитела, остав-шиеся в сыворотке, реагируют с эритроцитами В (Pacuzska и соавт. [181]). Иммунные кроличьи анти-В-антитела не реагируют с эритроцитами цис-АB.
В то же время адсорбция анти-В-сывороток человека эритроцитами кроли-ка полностью нейтрализует их активность по отношению к эритроцитам цис-АB. Важная деталь: антиген Н экспрессирован на эритроцитах цис-АB сильнее, чем на А2 и А2В. Сыворотки крови лиц цис-АB часто содержат анти-В-антитела (Daniels [87], Reid и соавт. [186]). Сыворотка крови женщины, генотип которой был АВ / О, имела нормально выраженный антиген В и антител анти-В не содер-жала (Pacuzska и соавт. [181]). В слюне лиц цис-АB выявлена нормальная кон-центрация вещества А, повышенная концентрация вещества Н. Субстанцию В определяли в следовых количествах (Pacuzska и соавт. [181]).
В ряде случаев лица цис-АB были описаны как А2В3. Экспрессия антигена А
у них была такой же, как у лиц А2, в то время как антиген В был выражен очень слабо. В одной французской семье, напротив, пробанд цис-АB имел фенотип А1В, хотя экспрессия антигена В была снижена (Salmon и соавт., 1984).
На присутствие аллеля цис-АB указывают несколько моментов. Во-первых, атипичное наследование группы крови у членов семьи. Во-вторых, антиген В экспрессирован слабее, чем у лиц АВ (имеющих генотип А / В). В-третьих, сы-воротка крови таких лиц реагирует с эритроцитами В.
По-видимому, цис-АB является результатом кроссинговера локусов А и В, в ре-зультате чего возникает гибридый ген, контролирующий А- и В-гликозилтранс фе-разную активность одновременно.
Геногеография
К настоящему времени накоплен обширный материал о распределении групп крови среди представителей различных рас и этнических групп (Ю.Г. Рычков и др. [10], А.К. Туманов и В.В. Томилин [60], Mourant и соавт. [165]). Ген О име-ет высокую частоту среди коренного населения Северной и Южной Америки, в некоторых регионах Африки, а также у австралийских аборигенов. Среди жи-телей Европы часто встречается ген А. Самая высокая частота гена В зафик-сирована среди монголоидов Сибири и Дальнего Востока и жителей Индии.
В северном полушарии по мере продвижения с запада на восток (из Европы в Азию) частота гена А снижается, а гена В увеличивается (Garratty и соавт. [109], Miyashita, Hasekura [155], Mourant и соавт. [165]).
Результаты исследований, проведенных в России и сопредельных странах, свидетельствуют о существенных различих в распределении генов АВО у пред-ставителей разных рас и этнических групп (А.С. Абдина [1, 2], А.Г. Башлай [4], Н.П. Вожегова и др. [8, 9], Ю.Г. Рычков и др. [10], Г.М. Давыдова [13], Г.А. Зайцева
и др. [29], Н.Н. Меркулова [41], Н.М. Михайлова, Н.И. Васильев [46], В.А. Мороков
[48, 50], Е.А. Хромова [63], В.Н. Шабалин и Л.Д. Серова [64]) (табл. 3.4).
68
Выделяют триаду, характерную для монголоидов: преобладание феноти-пов O(I) или B(III), Rh + и K–. Можно добавить четвертую особенность – Di a +. Вместе с тем неодинаковая частота групп крови не может служить дифферен-циальным критерием, отличающим представителей одной расы от другой. Вряд ли можно с точностью установить расовую принадлежность конкретного ин-дивида по сочетанию групповых антигенов его эритроцитов и рассматривать групповую принадлежность крови как признак европеоидности или монголоид-ности. В этом отношении набор групповых антигенов менее информативен, чем строение тела и цвет кожи. Сходное распределение групповых антигенов эри-троцитов было выявлено в популяциях, населяющих районы земного шара, уда-ленные друг от друга на многие тысячи километров. Так, частота групп крови АВО практически одинакова у французов и аборигенов Новой Гвинеи (Mourant
и соавт. [165]), англичан и хакасов (А.С. Абдина [1]). Mourant и соавт. [165], впервые составивший геногеографическую карту мира, не без удивления обра-тил на это внимание.
Несмотря на разную частоту групп крови у представителей отдельных рас и этнических групп, четко очерченное национальное своеобразие, специфических расовых или каких-либо этнических антигенов не обнаружено. Австралийский антиген, который, как вначале полагали, свойствен австралийским аборигенам, оказался фрагментом вируса гепатита, встречающегося и у европеоидов, и у монголоидов. Попытки установить дифференциальные расовые признаки с по-мощью молекулярно-генетических методов также ни к чему не привели (Olsson
и соавт. [174, 176–179]).
Таблица 3.4
Дата: 2019-02-24, просмотров: 513.