Широкие возможности воздействия на функции многих органов создают вещества, изменяющие (или имитирующие) влияние эфферентных (центробежных) нервов. Понимание действия этих веществ требует представлений о механизмах межклеточной передачи нервных сигналов, природе медиаторов и их рецепторов, типах эфферентных нервов и их влиянии на функции различных, в частности, внутренних органов.
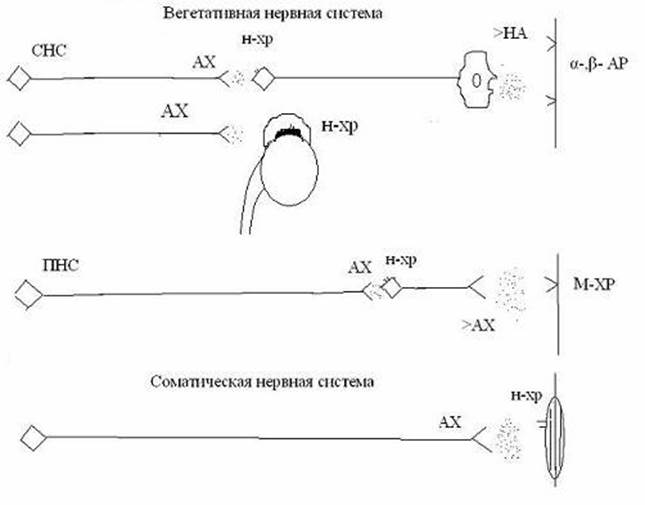
Эфферентная нервная система представлена соматическими и вегетативными нервами. Соматические нервы, управляющие произвольной (скелетной, поперечно-полосатой) мускулатурой. Вегетативные нервы иннервируют внутренние органы. Различают симпатические и парасимпатические нервы. Все вегетативные нервы имеют двухнейронное строение, состоят из пре- и постганглионарных нейронов. Медиатором всех преганглионарных вегетативных нейронов и соматических нервов является ацетилхолин. Эти нервы и образуемые ими синапсы называются холинергическими. В постсинаптической мембране таких синапсов (скелетных мышцах, нейронах симпатических и парасимпатических ганглиев, а так же хромаффинных клетках надпочечников и клетках синокаротидных телец) находится никотиночувствительные (Н-) –холинорецепторы (рецепторы-каналы). Их активация увеличивает проводимость постсинаптических мембран клеток для катионов, что ведет к возбуждению иннервируемых клеток.
Постганглионарные парасимпатические волокна иннервируют клетки круговой мышцы радужки и цилиарной мышцы глаза, мышечные и железистые клетки внутренних органов. Медиатором постганглионарных парасимпатических нервов является ацетилхолин, но в иннервируемых клетках (а также в потовых железах) находятся не никотиночувствительные, а мускариночувствительные (М-) холинорецепторы (метаботропные). Они делятся на подтипы: М1, М2, М3. Активация М1- и М3-ХР ведет к сокращению клеток гладких мышц и усилению секреторной деятельности железистых клеток. Активация М2-ХР в миокарде угнетает его деятельность.
Постганглионарные симпатические нейроны иннервируют мышечные, железистые клетки внутренних органов, клетки сосудов, радиальной мышцы радужки, мышц волосяных мешочков, потовые железы. Медиатором симпатических постганглионарных нервов является норадреналин. Соответствующие нервы и синапсы называются адренергическими Находящиеся в исполнительных органах адренорецепторы (альфа и бета) метаботропного типа. Одни из них – синаптические , обеспечивающие влияние симпатических нервов (альфа1, бета1), другие – внесинаптические, обеспечивающие влияние адреналина, гормона мозгового вещества надпочечника (альфа2, бета2). Активация α1-АР вызывает повышение концентрации в цитоплазме клеток концентрации ионов кальция. α2- АР запускают разные механизмы трансдукции и повышают тонус сосудов, усиливают агрегацию тромбоцитов, вызывают расслабление миоцитов желудка и кишечника. Активация всех видов бета-адренорецепторов вызывает повышение концентрации цАМФ в цитоплазме и активацию протеинкиназы А. Это приводит к изменению тонуса полых гладкомышечных органов, в том числе и сосудов, миокардиоцитов и
| Изменение функции органов при раздражении нервов (активации рецепторов) | ||
| Симпатических нервов адренорецепторов: α1 α2 β1 β2 | Органы (в скобках – тип имеющихся рецепторов) | Парасимпатических нервов холинорецепторов: М2, М3 |
| сокращение, мидриаз - расслабление | ГЛАЗ, мышцы: Расширяющая зрачок (α1) Суживающая зрачок (М3) Цилиарная (М3) | - сокращение, миоз сокращение |
| - усил.,густая слюна усиление | ЖЕЛЕЗЫ, секреция Слезные (М3) Слюнные (М3) Потовые (М3) | усиление усилен., жидкая слюна - |
| снижается снижается | БРОНХИ тонус мышц (М3 β2) секреция желез (М3) | повышается повышается |
| повышается Повышается ускоряется повышается | СЕРДЦЕ частота сокращений (β1 М2) сила сокращений(β1 α1(желудочки), М2) проводимость (β1 М2) возбудимость (β1 М2) | снижается Уменьшается замедляется понижается |
| суживаются суживаются расширяются суживаются | СОСУДЫ кожи (α1, α2) мезентериальные (α1, α2) скелетных мышц (β2) сердца, мозга | - - расширяются расширяются |
| сокращение | Миоциты трабекул селезенки (α1) | - |
| усиление агрегации | Тромбоциты (α2) | |
| торможение выброса медиаторов немедленной аллергии (дегрануляции) | Базофилы, тучные клетки (β2) | |
| ослабляется повышается усиливается (добавочных) | ЖЕЛУДОК, КИШЕЧНИК Перистальтика (М3 α2, β1(кишечн.)) тонус сфинктеров (α1) секреция желез желудка | усиливается снижается усиливается |
| расслабляются | ЖЕЛЧНЫЙ ПУЗЫРЬ Протоки (М3) | сокращаются |
| уменьшение (α2) увеличение (β2) секреции инсулина | Клетки Лангерганса поджелудочной железы | |
| гликогенолиз, гипергликемия | Гепатоциты (α1, β2) | |
| усиление липолиза (β3) угнетение липолиза (α2) | Жировые клетки (β3, α2) | |
| увеличение высвобождения ренина | Юкстагломерулярные клетки (β1) | |
Продолжение таблицы 2
| Изменение функции органов при раздражении нервов (активации рецепторов) | |||
| сокращение | МОЧЕТОЧНИКИ (α1) | - | |
| расслабление сокращение | МОЧЕВОЙ ПУЗЫРЬ Детрузор (М3) Сфинктер (α1) | сокращение расслабление | |
| сокращение(16-37,40нед.) Расслабление | БЕРЕМЕННАЯ МАТКА Тело, шейка α1 (β2) | - сокращение | |
| уменьшение импульсного высвобождения медиатора | Терминали адренергических аксонов (α2) | ||
секреции желез (см. таблицу).
Влияние холин- и адренергических нервов на иннервируемые органы опосредовано синапсами – местами наиболее тесного контакта иннервирующей и иннервируемой клеток. В зависимости от выделяемого нервными окончаниями медиатора различают холинергические и адренергические синапсы. Любой синапс состоит из трех частей: пресинапса (окончание соответствующего нерва), постсинапса (участок цитоплазматической мембраны иннервируемой клетки) и узкой заполненной внеклеточной жидкостью синаптической щели. В любом синапсе протекают 6 процессов: 1.синтез медиатора; 2.накопление (депонирование) медиатора в пузырьках (везикулах), транспорт везикул в нервное окончание – пресинапс и фиксация множества везикул вблизи пресинаптической мембраны; 3.вызванное пришедшим в окончание нерва потенциалом действия (импульсное) высвобождение медиатора, диффузия медиатора в жидкости синаптической щели; 4.активация медиатором постсинаптических рецепторов (рецепция медиатора); 5.вызыванное медиатором изменение белковой структуры рецептора, приводящее к повышению ионной проводимости постсинаптической мембраны, либо к активации мембранных ферментов, образованию вторичных посредников и активации протеинкиназ и изменению активности иннервируемой клетки (трансдукция); 6.удаление медиатора из синаптической щели с помощью ферментов или транспортеров (инактивация медиатора).
Вещества, изменяющие синаптическую передачу, могут воздействовать на процессы в пресинапсе (синтез, депонирование, импульсное высвобождение медиатора), в постсинапсе (рецепция, трансдукция) или на инактивацию медиатора.
Ацетилхолин (АХ) синтезируется из холина и уксусной кислоты и депонируется в синаптических пузырьках и вневезикулярных депо. Потенциал действия, достигая терминали аксона холинергического нейрона и вызывая деполяризацию ее мембраны, вызывает поступление через п/з Са-каналы Са2+ внутрь терминали и высвобождение содержимого синаптических пузырьков в синаптическую щель (экзоцитоз). Высвобожденные нервным импульсом молекулы АХ диффундируют в пространстве синаптической щели, соударяются с поверхностью постсинаптической мембраны, активируя встроенные в нее холинорецепторы. В синаптической щели присутствует фермент ацетилхолинэстераза, которая разрушает АХ (гидролизует до холина и уксусной кислоты). Понижение концентрации АХ в синапсе ведет к диссоциации комплексов АХ-ХР, после чего ХР переходят в исходное состояние и синапс оказывается способным к восприятию следующего импульса. Везамикол блокирует вход ацетилхолина в везикулы. Ионы Mg2+ и аминогликозиды препятствуют входу Са2+ в нервное окончание через п/з кальциевые каналы (аминогликозиды могут нарушать нервно-мышечную передачу). Ботулиновый токсин вызывает протеолиз синаптобревина (белок мембраны везикул, который взаимодействует с белками пресинаптической мембраны) и поэтому препятствует встраиванию везикул в пресинаптическую мембрану. Таким образом уменьшается выделение ацетилхолина из холинергического окончания. При ботулизме нарушается нервно-мышечная передача; в тяжелых случаях возможен паралич дыхательных мышц. 4-Аминопиридин блокирует К+-каналы пресинаптической мембраны. Это замедляет реполяризацию потециалов действия, увеличивает их длительность, усиливает вхождение Са2+ в пресинапс и высвобождению ацетилхолина. 4-Аминопиридин облегчает нервно-мышечную передачу.
Норадреналин синтезируется из фенилаланина в несколько этапов. Последовательно, с участием ряда ферментов образуется тирозин, диоксифенилаланин (ДОФА), дофамин (ДА), норадреналин. До образования дофамина процесс осуществляется в цитоплазме адренергических нейронов, образующийся ДА активно транспортируется в везикулы, где он превращается в норадреналин. Синтезированный норадреналин запасается в везикулах и вневезикулярных депо. Последний не высвобождается нервным импульсом и может служить источником пополнения запасов медиатора для вновь образующихся везикул. При чрезмерном синтезе медиатора и насыщении всех мест его депонирования избыток образующегося норадреналина разрушается ферментом моноаминоксидазой. Достигающие терминалей адренергических аксонов нервные импульсы, вызывая вхождение ионов Са2+, высвобождают запасенный в везикулах медиатор механизмом экзоцитоза. Высвобожденный из синаптических пузырьков норадреналин, диффундируя в синаптической щели, достигает поверхности постсинаптической мембраны, где часть его инактивируется катехол-О-метилтрансферазой (КОМТ), но большая часть молекул медиатора взаимодействует с альфа- и бета-адренорецепторами (АР), что сопровождается их активацией. Около 80 % высвобождаемого нервным импульсом норадреналина возвращается в пресинапс с помощью находящейся в пресинаптической мембране транслоказы, пополняя вневезикулярные запасы норадреналина, а затем редепонируя в везикулах.
Дата: 2019-03-05, просмотров: 459.