В этом руководстве речь пойдет только об эндок-ринопатиях, затрагивающих половую дифференциров-ку, созревание и функцию гонад как желез внутренней секреции. Для организма подростка гонады — основные дирижеры пубертата, то есть их функция и состоит в данный период не в размножении инди-
вида, но в завершении его половой дифференци-ровки.
До определённой стадии эмбриогенеза каждый индивид представляет собой соматического «андрогина» с недифференцированным бипотентным гонадным зачатком (гонадным валиком), содержащим первичные половые хорды и мигрировавшие из стенки желточного мешка примордиальные половые клетки, способные к альтернативной дифференцировке.
Эта структура формируется на 4-й неделе внутриутробной жизни. Она содержит корковое и мозговое вещество. На 7-8-й неделе происходит дивергенция и зачатки дают либо мужскую, либо женскую гонаду. В мужском варианте преимущественное развитие получают мозговые, а в женском — корковые структуры половых валиков. При этом хорды превращаются либо в семенные канальцы, либо в овариальные фолликулы; ме-зенхимные клетки формируют либо клетки Лейдига, либо текальные и стромальные элементы яичника, а примордиальные половые клетки превращаются в сперма-тогонии или овогонии, соответственно.
Каждый плод располагает вольфовым (мезонеф-ральным) протоком, оформляющимся к 25-30-му дню беременности, и мюллеровым (парамезонефральным) протоком, формирующимся к 44-48-му дню (Кнорре А.Г., 1959, Улумбеков Э.Г., Челышев Ю.А., 1997). Вольфов проток может дифференцироваться в эпидидимис, семявыносящий проток, семенные пузырьки и эякуля-торные протоки. Мюллеров может дать фаллопиевы трубы, матку и верхюю часть влагалища.
Наконец, имеются универсальные зачатки наружных гениталий—урогенитальный синус, мочеполовые бугорок, складки и валики. Эти структуры можно преобразовать либо в пенис и мошонку, либо в клитор и половые губы.
Напомним, что мужской пол развивается только при наличии Y-хромосомы, а еще точнее — её SRY-гена, если правильно реализуется определённая цепь эмбри-огенетических событий. При отсутствии Y-хромосомы действует спонтанная тенденция к развитию всех бивалентных эмбриональных зачатков по женскому типу, которая не требует особых гормональных активаторов. При отсутствии гонад у плода вольфов проток регрессирует, а мюллеров дифференцируется по женскому типу. При кариотипе 45ХО (синдром Шерешевского-Терне-ра) формируется яичник, хотя последующее развитие в нем фолликулов нарушено, и для его полноценности требуется кариотип 46ХХ.
Развитие мужского пола при кариотипе, лишенном Y хромосомы, зафиксировано только в редчайших эк-сквизитных случаях переноса SRY гена из Y хромо-
315
|
|
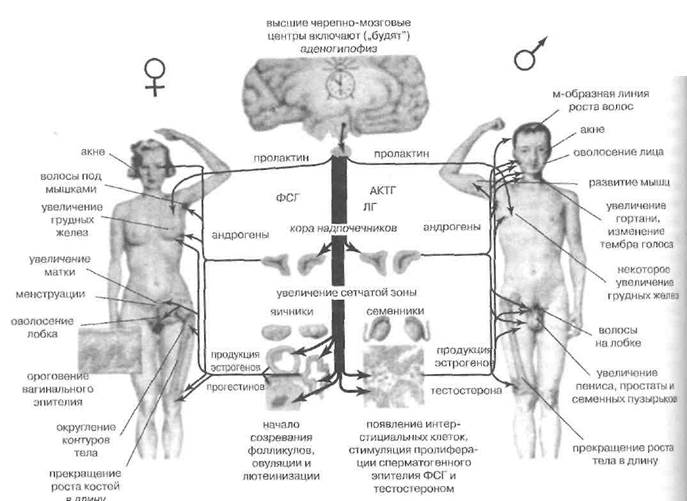
Рис. 88. Нормальная половая дифференцировка и половое созревание у человека (по Ф. Неттеру, США)
сомы — в Х-хромосому из за неправильного кроссин-говера при гаметогенезе. Таких наблюдений около 50. При этом кариотип 46ХХ, а фенотип мужской. Больные низкого роста, имеют нормальный пенис, маленькие плотные яички, в которых, однако, практически нет сперматозоидов. Уровень гонадотропинов повышен, тестостерона — снижен, эстрогенов — выше нормы.
Целостный ход процесса половой дифференцировки и пубертата у подростков отражен на рис. 88.
Критической в процессах развития первичных мужских половых признаков является 8-я неделя внутриутробного развития.
Чтобы произошла полная реализация программы маскулинизации, ген SRY из семейства ДНК-регулятор-ных генов Sox, находящийся в коротком плече Y хромосомы, близ центромера, вступает в действие на 6-7-й неделе эмбрионального развития, запуская ряд генов, локализованных в других хромосомах и обеспечивающих программу синтеза тестостерона у плода, выработку фермента Д4-5а-редуктазы, необходимой
316
для конверсии этого андрогена (рис. 56) в дигидроте- стостерон (эти гены находятся в аутосомах) и синтез пептида, ингибирующего развитие мюллерова протока и тканевых рецепторов андрогенов (эти гены присутствуют в Х-хромосоме). Важное значение имеет ген SRA1 семейства Sox, локализованный в хромосоме 17, кодирующий дополнительные факторы, опосредующие часть эффектов гена SRY.
Белковые продукты, синтез которых запускается геном SRY, представляют собой «мужские антигены», ис пользуемые в иммунологическом тестировании ис тинного пола. Мужской антиген H-Y, он же — фактор, определяющий развитие тестикул (ФРТ), служит иммунологическим маркером мужского пола, отсутствует при отсутствии Y хромосомы и при делеции ее SRY-участ-ка, в то же время при наличии более чем одной Y-xpo-мосомы его концентрации в крови кратно повышаются.
При мутациях в 17-й хромосоме и отсутствии функций гена SRA1 действие ФРТ не полностью эффективно. Происходит развитие плодов с кариотипом 46XY по женскому типу (реверсия пола) или развивается врож-
 дённая камптомелическая дисплазия, при которой большинство больных-мужчин имеет женский фенотип. Взаимоотношения этого гена с расположенным там же геном хорионического гонадотропина не уточнены. Процесс первичной мужской половой дифференци-ровки занимает 8-14-ю недели эмбриогенеза. До 50-го дня гонады индифферентны. Действие продуктов гена SRY (и, возможно, SRA1) ведет на 7-8-й неделе к диф-ференцировке первых клеток Лейдига и клеток Серто-ли в зачатке гонады. Клетки Лейдига начинают продукцию фетального тестостерона, их деятельность стимулируется плацентарным хорионическим гона-д отропином.
дённая камптомелическая дисплазия, при которой большинство больных-мужчин имеет женский фенотип. Взаимоотношения этого гена с расположенным там же геном хорионического гонадотропина не уточнены. Процесс первичной мужской половой дифференци-ровки занимает 8-14-ю недели эмбриогенеза. До 50-го дня гонады индифферентны. Действие продуктов гена SRY (и, возможно, SRA1) ведет на 7-8-й неделе к диф-ференцировке первых клеток Лейдига и клеток Серто-ли в зачатке гонады. Клетки Лейдига начинают продукцию фетального тестостерона, их деятельность стимулируется плацентарным хорионическим гона-д отропином.
Фетальный тестостерон направляет развитие зародыша по мужскому варианту. Он способствует на 7-10-й неделе развития преобразованию вольфова прото-в эпидидимис, семенные пузырьки, эякуляторные протоки и семявыносящий проток. В тканях, под воздействием экспрессируемой при наличии гена SRY А4-5а-редуктазы, тестостерон переходит в дигидроте- стостерон, необходимый для преобразования общего зачатка наружных гениталий по мужскому типу — в мошонку и пенис, а также для развития простаты.
Эти процессы завершаются к 14 неделям беременности, хотя нисхождение яичек в мошонку и рост пениса продолжаются и в дальнейшем. В то же время, с начала восьмой недели первые клетки Сертоли вырабатывают локальный фетальный гормон—высокомолекулярный гликопротеид, известный под названием мюллеров тормозный фактор (антимюллеров ин-гибирующий пептид). Этот регулятор обеспечивает быструю инволюцию мюллерова протока, к началу 9-й недели беременности, но продолжает выделяться у плода мужского пола до момента родов и даже в раннем постнатальном периоде. Его гомолог выделяется яичками и в последующем (см. ниже).
Итак, продукция тестостерона и антимюллеро-ва ингибирующего пептида в гонадах плода предопределяет на втором-третьем месяцах внутриутробного развития формирование внутренних половых органов по мужскому типу. Метаболит андрогенов 5 а-дигидротестостерон, не превращаемый в эст рогены, контролирует маскулинный тип формирования наружных гениталий на 3-4-м месяце фетогенеза соматический пол).
В то же время, значительная часть тестостерона у плода любого пола и в плаценте метаболизируется в зстрогены. При наличии Y-хромосомы и высокой продукции тестостерона обеспечивается достаточное содержание эстрогенных метаболитов, подавляющее ормирование обратной связи между продукцией лю-
теинизирующего гормона (ЛГ) и эстрогенов в развивающемся мозге. Это ведет к установлению мужского, нециклического типа гипоталамо-гипофизарной регуляции гонадотропных функций и обеспечивает на 4-6-м месяце фетогенеза мужской тип дифференциров-ки головного мозга и гипоталамо-гипофизарного ней-росекреторного аппарата, что лежит в основе психофизиологической маскулинизации в последующей жизни.
Созревание центров секреции гонадотропинов идет под контролем эстрогенов, полученных из тесто стерона, а центров, определяющих половое влечение —под совместным контролем андрогенов и их ароматических эстрогенных производных. Центры, ответственные за поддержание поведения, соответствующего избранной половой роли, у мужчин организуются под влиянием только андрогенов. Организация трех этих мозговых центров идет последовательно и частично перекрывается.
Циклический тип секреции гонадотропинов мозга и высокая чувствительность механизма обратной связи между концентрацией эстрогенов и продукцией ЛГ обнаружены у истинно гомосексуальных мужчин, но отсутствуют у гетеро- и бисексуалов. Таким образом, ис тинный первичный гомосексуализм не должен трактоваться как чисто психосоциальное явление, так как он является болезненным нарушением полового развития мозга (Г. Дёрнер и соавт., 1987), а психолого-поведенческие проявления носят по отношению к этому вторичный характер.
Значительную роль в опосредовании половой диф-ференцировки мозга под влиянием андрогенов играют норадренергические и серотонинергические механизмы. Установлено снижение скорости захвата серотонина постсинаптическими нервными окончаниями и тромбоцитами лиц с парафилиями и транссексуальными нарушениями поведения (Ткаченко А.А. с соавт., 1995). По данным И. Лехтинена с соавт. (1971), блокаторы синтеза серотонина феминизируют половое поведение самцов грызунов, а ингибиторы моноаминооксидазы, способствующие накоплению катехоламинов и серотонина, маскулинизируют поведение самок.
В постнатальном онтогенезе, в период полового созревания, активизация стероидогенной функции семенников, выходящих из-под ингибирующего действия эпифиза, ведет к приросту продукции андрогенов — тестостерона и дигидротестостерона — и к маскулинизации внешнего облика подростков, сопровождаемому развитием мужских вторичных половых признаков, гиперплазией тестикул и наружных гениталий, началом полноценного сперматогенеза, который про-
317
должается всю жизнь. По крайней мере, зафиксированы случаи рождения полноценных детей у глубочайших старцев, в том числе лиц более чем 100-летнего возра-ста. Хрестоматийный пример — рождение философа Конфуция, отцу которого к тому моменту перевалило далеко за 80.
В норме половое созревание начинается у мальчиков в 10-12 лет. Это происходит благодаря первичным изменениям в сервомеханизмах, определяющих взаимоотношения тестикул и гипоталамо-гипофизарного аппарата. Перед пубертатом гипоталамус и гипофиз очень чувствительны к ингибирующему влиянию тестосте рона на продукцию гонадолиберина и гонадотропи-нов. Начало пубертата знаменуется понижением этой чувствительности. Появляются ночные всплески гона-долибериновой и лютропиновой секреции. Затем они распространяются и на дневное время. Наконец происходит переустановка гипоталамуса на новый уровень чувствительности к андрогенам. Секреция гонадолиберина и чувствительность гонадотропов к гонадолиберину возрастают. Это ведет к стимуляции ЛГ продукции те стостерона в яичках и к их гиперплазии. Андрогены обеспечивают формирование признаков возмужания, в основном — путём дифференцированной стимуляции ростовых и анаболических процессов. При этом до чет верти всей задержки азота, обусловленной андро-генами и связанной у подростков с активизацией синтеза нуклеиновых кислот и белков, приходится на пролиферативно-гипертрофические прцессы в гонадах и гениталиях.
Остальная задержка азота связана с ростом скелета и мышечной массы, ростом волос и гипертрофией сальных желез, ростом и изменением конфигурации гортани и, наконец—активизацией эритропоэза. Особенно чувствительны к андрогенам мышцы плечевого пояса и груди, в связи с чем маскулинизируются очертания фигуры. Весь процесс пубертата занимает около 5 лет, после чего дополнительная андрогенизация у молодых людей уже не усиливает выраженности половых признаков. Правда, отдельные проявления маскулинности — например, мышечная масса и степень оволосения груди, — могут нарастать на протяжении более длительного срока. В отсутствие гена SRY, все вышеописанные события не происходят и формируется соматический и психический женский пол, причём примордиальная гонада превращается в яичник.
В диапазоне между 50-м и 90-м днём беременности формируются и успевают достичь стадии ооцитов I порядка женские гаметы. Они обрывают своё развитие в профазе первого мейотического деления и приобретают оболочку из фолликулярных клеток, образуя
318
примордиальные фолликулы, которых к 7-му месяцу внутриутробной жизни насчитывается до 10 млн, а к рождению их число падает до 2 млн. Вольфов проток регрессирует к 10 неделям беременности, мюллеров дифференцируется во внутренние гениталии, а общий зачаток наружных гениталий после 11-й недели без влияния феталъных андрогенов даёт наружные женские половые органы. В дальнейшем развитии женских га- мет следует период покоя. За его время число ооцитов сократится до 400 000.
В постнаталъной жизни единственным эндогенным источником андрогенов в организме здоровых женщин служит кора надпочечников. В период пубертата активизация внутрисекреторной деятельности яичников, продуцирующих женские по ловые стероиды (эстрогены и прогестерон) осуще-ствляемая под контролем гипоталамуса и гипофиза, в соответствии с заложенным в фетальный период циклическим паттерном, приводит к бурному развитию вторичных женских половых признаков и началу месячных. Период покоя в овогенезе заканчивается, на пике содержания ЛГ завершается первое мейотическое деление, образуются и циклически выходят в половые пути полноценные яйцеклетки. Только немногим более 400 из них будет использовано за всю предстоящую жизнь половозрелой женщины.
Таким образом, если мужчина, при длящемся много десятилетий процессе самообновления сперматоген-ного эпителия тестикул, имеет буквально миллиарды шансов стать отцом, даже у женщин идеального здо-ровья это число шансов ограничено всего лишь сотня-ми. Авторам кажется, что это — факт, полезный для эффективной санитарно-просветительной работы юве-ногинеколога (Балахонов А.В., 1999,2000).
Следует отметить, что большое количество сперматозоидов уравновешивается тем, что каждый из них намного более уязвим для неблагоприятных влияний, чем отдельно взятая яйцеклетка. Поэтому частота женского и мужского бесплодия практически одинакова, несмотря на разительные отличия в количественных результатах ово- и сперматогенеза.
Отметим также, что яичник, в отличие от яичка, очень плюрипотентная эндокринная железа. Помимо половых стероидов, различные клетки яичника являются источником еще более чем 30 белковых гормонов и паракринных регуляторов (Окленд Дж. с соавт., 1992). В минорных количествах все они обеспечивают нормальную регуляцию женских половых функций. Однако при патологии именно яичник может служить эктопическим источником избытка ряда гормонов.
 Асинхрония рассмотренных выше процессов формирования половых признаков и их зависимость от таких метаболических факторов, как ароматизация половых стероидов и их рецепция тканями, создают почву для множества врождённых и приобретённых рассогласований и нарушений соответствия хромосомного, гонадного, нейроэндокринного, психического и соматического пола. В большинстве случаев эти нарушения проявляются именно у подростков. Наиболее важные из них рассмотрены ниже.
Асинхрония рассмотренных выше процессов формирования половых признаков и их зависимость от таких метаболических факторов, как ароматизация половых стероидов и их рецепция тканями, создают почву для множества врождённых и приобретённых рассогласований и нарушений соответствия хромосомного, гонадного, нейроэндокринного, психического и соматического пола. В большинстве случаев эти нарушения проявляются именно у подростков. Наиболее важные из них рассмотрены ниже.
ИСТИННЫЙ ГЕРМАФРОДИТИЗМ
Определение. Присутствие у индивида ткани как I яичек, так и яичников называется истинным гермаф- \родитизмом.
Распространенность. Истинный гермафроди-
тизм издавна вызывал пристальное внимание медиков,
поскольку идея «промежуточного существа» глубоко
укоренилась в культуре и философии. О гермафродитах
мы можем прочесть в античных мифах, описывающих
прототипа, давшего название этому синдрому—сына
Гермеса и Афродиты, слившегося в единое существо
с нимфой Сольмацией, а также в философских трудах
B.C. Соловьёва, считавшего стремление к андрогин-
ности направлением эволюции человека.
Один из интереснейших случаев истинного гермафродитизма, когда гермафродит имел нормальных детей в браке, описан выдающимся немецким поэтом доктором медицины И.В. Гёте.
Но наиболее эксквизитное описание поистине уникального случая истинного гермафродитизма принадлежит Рудольфу Вирхову и соавторам. Ими был описан истинный гермафродит, при рождении названный Катарина Гоман и воспитанный как девочка. В 16 лет этот подросток посчитал себя мужчиной и начал половую жизнь с женщинами. В 20 лет у него появились менструации и гинекомастия. Психофизиологическая ориентация гермафродита изменилась, и он начал вести половую жизнь с мужчинами, как нормальная женщина, но детей не имел. В 42 года менструации прекратились. Тогда этот замечательный исторический персонаж еще раз изменил стереотип своего сексуального поведения и, переменив имя Катарина на Карл Гоман, женился и имел в браке здорового сына. Обследовав Катарину-Карла, Р. Вирхов обнаружил живые сперматозоиды, а правильные менструации говорили о наличии функционального яичника. Со времён Вирхова подобных случаев больше не описано, хотя имеются свидетельства о более чем четырёх сотнях менее ярких наблюдений истинного гермафродитизма.
Этиология и патогенез. Нарушение является результатом мозаичных генетических транслокаций. Патогенез истинного гермафродитизма связан с судьбой гена SRY. Кариотип истинных гермафродитов — в 80% случаев 46ХХ, в большей части оставшихся наблюдений — мозаичный 46XX/XY и лишь изредка — 46XY. Большинство из больных имеют мозаицизм с переносом в части клеток SRY-гена на Х-хромосому или на аутосому. У некоторых гермафродитов с мозаицизмом Н. Йоссо с соавт. (1965) обнаружили признаки, указывающие на двойное оплодотворение при их зачатии.
Клетки из овариальной части гонадной ткани больных при мозаицизме не имеют антигена H-Y, а из тес-тикулярной части—располагают этим маркером. Доза антигена понижена из-за инактивации несущих эктопический SRY-ген хромосом в части соматических клеток, по случайному принципу.
Есть точка зрения, что истинный гермафродитизм, как и синдром «кроссинговерных мужчин 46ХХ», являются промежуточным и крайним проявлениями одного и того же заболевания. По крайней мере, при семейных случаях истинного гермафродитизма описана родословная, где брат и дядя пробанда — истинного гермафродита имели этот синдром.
Клиника. У истинного гермафродита могут быть один яичник и один семенник или смешанная овотес-тикула и яичник (в более чем 60% наблюдений). Реже бывают двусторонние овотестикулы (более 20%), а самое редкое сочетание — овотестикула и яичко (10%) При этом обычно в разнополых гонадах или разнополых частях овотестикул имеются гаметы разных полов. Часть истинных гермафродитов формирует гаметы лишь одного пола. Не более половины из них имеют зрелые яйцеклетки, а достаточное для мужской фертильности количество полноценных сперматозоидов образуют лишь отдельные истинные гермафродиты.
Строение наружных половых органов у истинных гермафродитов промежуточное, около 7% имеют наружные гениталии женского типа, в 12% наблюдений присутствует пенис и уретра. Однако в 75% случаев истинные гермафродиты в детстве и юности воспитываются и осознают себя в социально-половом отношении как мужчины (Императо-Мак-ДжинлиДж., 1987).
При истинном гермафродитизме не описано случаев самооплодотворения, поскольку к периоду полового созревания сохраняется формирование зрелых гамет лишь одного типа, почти всегда — яйцеклеток (Либерман Л.Л., 1966). В пубертатный период иногда происходит смешанная вирилизация-
319
феминизация. Чаще преобладает влияние эстрогенов. До 80 % истинных гермафродитов имеют гинекомастию, почти 50 % менструируют (при наличии уретры и гипоспадии — в форме периодической гематурии).
Прогноз. Истинные гермафродиты могут иметь детей. Зафиксировано много случаев беременности и успешных родов у истинных гермафродитов, ведущих половую жизнь как женщины, при условии удаления остатков тестикул и коррекции наружных гениталий. Отдельные истинные гермафродиты, ведущие половую жизнь, как мужчины, также являются фертильными.
Истинные гермафродиты имеют нормальный интеллект и физическое развитие, но рост их часто — ниже среднего. Продукция гонадотропинов не понижена, а часто — увеличена.
Истинный гермафродитизм предрасполагает к возникновению злокачественных опухолей из половых клеток — гонадобластом, которые поражают овотестику-лу или яичко в 3% случаев.
ЛОЖНЫЙ ГЕРМАФРОДИТИЗМ
Определение. Ложный гермафродитизм (синоним — псевдогермафродитизм) — синдром, при котором наличествуют гонады одного пола и половая принадлежность имеющихся гонад соответствует кариотипу (46ХУ — при мужском и 46ХХ — при женском псевдогермафродитизме). Однако формирование других половых органов и вторичных половых признаков характеризуется при данном синдроме интерсексуальностью.
Мужской псевдогермафродитизм — результат разнообразных нарушений в эмбриогенезе гонад, продукции, метаболизме и рецепции андрогенов. Он характеризуется наличием парных семенников и неполной маскулинизацией (могут быть гипоспадия, микрофаллия, плохо развитая мошонка).
Женский псевдогермафродитизм — наиболее часто является результатом появления эндогенных или экзогенных андрогенов в критический период от 8-й до 12-й недель киематогенеза, когда плод с кариотипом 46ХХ чувствителен к их морфогенетическому влиянию. У больных имеются яичники, но фенотип, как правило, мужской. Это вызывается врождённой гиперплазией коры надпочечников, арренобластомой во время беременности у матери или приемом матерью андрогенов и ыв первом триместре беременности.
Псевдогермафродитизм распространен намного шире, чем гермафродитизм истинный. Так как перемена психолого-поведенческих половых стереотипов
320
у детей после 2 лет происходит с трудом, важной задачей является диагностика истинного пола и выбор желаемого пола с коррекцией проявлений псевдогермафродитизма по возможности в раннем детском, а не в подростковом возрасте.
Дата: 2019-02-02, просмотров: 420.