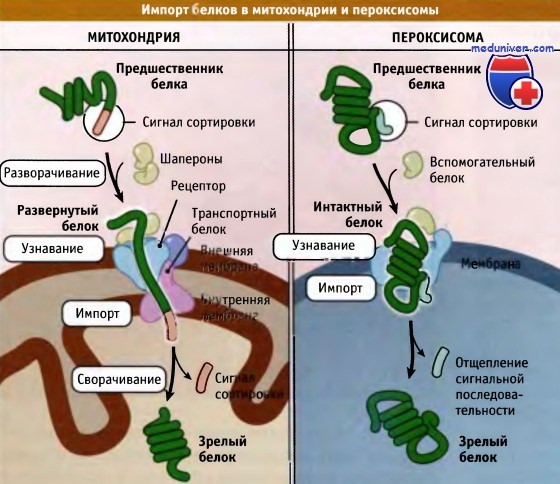
Для всех органелл, окруженных мембранами, существует одна общая проблема: каким образом получить белок, который образуется в цитозоле? При этом белок должен быть локализован либо в мембране, либо пройти через нее. Прохождение гидрофильного белка через гидрофобную мембрану невозможно из-за существования значительных энергетических препятствий. Каждая органелла выработала собственное решение этой общей проблемы. В ее мембране присутствуют белковые гидрофильные поры, через которые и осуществляется транспорт, так что белку не нужно взаимодействовать с гидрофобной мембраной. Строение самой поры и природа взаимодействия с транспортируемым белком зависят от органеллы. Ядерные поры представляют собой массивные структуры со сложным транспортным аппаратом, который определяет, какие белки должны транспортироваться в ядро, а какие выходить из него. Белки накапливаются на одной стороне оболочки, проходят через пору и выходят с другой стороны. Пора проходит через обе мембраны ядерной оболочки, и созревший белок транспортируется через всю пору. В наружной и внутренней мембранах таких органелл, как митохондрии и хлоропласты, содержатся белки, функция которых заключается в транспорте в органеллу импортируемых белков. Экспорта белков из этих органелл не происходит. Белки образуются на рибосомах в цитоплазме и выходят в цитозоль. В их структуре находятся специфические последовательности, которые взаимодействуют с рецепторами, находящимися в мембране органеллы (импорт белков в митохондрии начинается с узнавания сигнальной последовательности на внешней мембране). На рисунке ниже показано, что проходящий через мембрану канал очень узкий, и чтобы пройти через него, транспортируемый белок должен вначале развернуться, а на другом конце приобрести зрелую конфигурацию. Это требует активного участия дополнительных белков — шаперонов, контролирующих сворачивание белка (способность к свертыванию и развертыванию белковых структур является характерной особенностью всех клеток).
Опишите молекулярные и клеточные механизмы процессов РНК-интерференции (РНКи) в клетках эукариот. Использование РНКи для изменения активности генов и изучения функциональной роли кодируемых ими белков.
Принцип РНК-интерференции
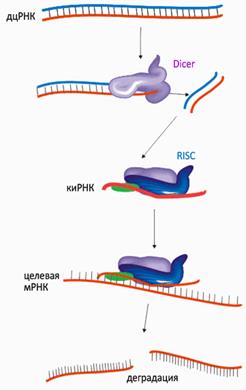 Сегодня изучение малых регуляторных РНК является одной из наиболее бурно развивающихся областей молекулярной биологии. Обнаружено, что все короткие РНК выполняют свои функции на основе явления, названного РНК-интерференцией (суть этого феномена заключается в подавлении экспрессии гена на стадии транскрипции или трансляции при активном участии малых молекул РНК). Очень схематично механизм РНК-интерференции показан на врезке «Основы РНК-интерференции».
Сегодня изучение малых регуляторных РНК является одной из наиболее бурно развивающихся областей молекулярной биологии. Обнаружено, что все короткие РНК выполняют свои функции на основе явления, названного РНК-интерференцией (суть этого феномена заключается в подавлении экспрессии гена на стадии транскрипции или трансляции при активном участии малых молекул РНК). Очень схематично механизм РНК-интерференции показан на врезке «Основы РНК-интерференции».
Двуцепочечные молекулы РНК (дцРНК) нехарактерны для нормальных клеток, но они являются обязательным этапом жизненного цикла многих вирусов . Специальный белок Dicer, обнаружив в клетке дцРНК, «режет» её на небольшие фрагменты. Антисмысловая цепь такого фрагмента, которую уже можно называть короткой интерферирующей РНК (киРНК, от siRNA — small interference RNA), связывается комплексом белков под названием RISC (RNA-induced silencing complex), центральный элемент которого — эндонуклеаза семейства Argonaute. Связывание с киРНК активирует RISC и запускает в клетке поиск молекул ДНК и РНК, комплементарных «шаблонной» киРНК. Судьба таких молекул — быть уничтоженными или инактивированными комплексом RISC. Подытоживая, короткие «обрезки» чужеродной (в том числе, введённой намеренно) двуцепочечной РНК служат «шаблоном» для широкомасштабного поиска и уничтожения комплементарных мРНК (а это эквивалентно подавлению экспрессии соответствующего гена), — причем, не только в одной клетке, но и в соседних. Для многих организмов — простейших, моллюсков, червей, насекомых, растений — этот феномен является одним из основных способов иммунной защиты против инфекций.
Дата: 2019-05-28, просмотров: 372.