У эукариотических организмов механизм регуляции транскрипции гораздо более сложен. В результате клонирования и секвенирования генов эукариот обнаружены специфические последовательности, принимающие участие в транскрипции и трансляции.
Для эукариотической клетки характерно:
1. Наличие интронов и экзонов в молекуле ДНК.
2. Созревание и-РНК - вырезание интронов и сшивка экзонов.
3. Наличие регуляторных элементов, регулирующих транскрипцию, таких как:
a) промоторы - 3 вида, на каждый из которых садится специфическая полимераза.
РНК-полимераза I - реплицирует рибосомные гены,
РНК-полимераза II - структурные гены белков,
РНК-полимераза III - гены, кодирующие небольшие РНК.
Промоторы РНК-полимеразы I и II находятся перед участком инициации транскрипции, промотор РНК-полимеразы III - в рамках структурного гена;
b) модуляторы - последовательности ДНК, усиливающие уровень транскрипции;
c) (энхансеры) усилители - последовательности, усиливающие уровень транскрипции и действующие независимо от своего положения относительно кодирующей части гена и состояния начальной точки синтеза РНК;
d) терминаторы - специфические последовательности, прекращающие и трансляцию, и транскрипцию.
Существенную роль в регуляции транскрипции у эукариот, помимо опосредованной взаимодействием между ДНК и белками, играют также белок-белковые взаимодействия.
Несмотря на индивидуальность набора регуляторных элементов у структурных генов эукариот, каждый из них имеет промоторный участок (ТАТА-бокс, или бокс Хогнесса) из восьми нуклеотидов, включающий последовательность TATA; последовательность ССААТ (САТ-бокс); участок из повторяющихся динуклеотидов GC (GC-бокс).
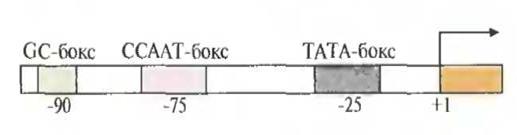
Транскрипция структурного гена эукариот начинается со связывания с ТАТА-боксом фактора транскрипции, который представляет собой комплекс по крайней мере из 14 белков. Затем с ним и участками ДНК, примыкающими к ТАТА-боксу, связываются другие факторы транскрипции, и, наконец, со всем этим транскрипционным комплексом связывается РНК-полимераза II. Затем при участии дополнительных факторов происходит инициация транскрипции в точке +1. Если последовательность TATA отсутствует или существенно изменена, то транскрипция структурного гена становится невозможной.
Процессинг м- РНК
Кэпирование представляет собой присоединение к 5'-концу транскрипта 7-метилгуанозина через необычный для РНК 5',5'-трифосфатный мостик, а также метилирование остатков рибозы двух первых нуклеотидов. Процесс кэпирования происходит во время синтеза молекулы пре-мРНК. Кэпирование защищает 5'-конец первичного транскрипта от действия рибонуклеаз, специфически разрезающих фосфодиэфирные связи в направлении 5’→3'.
Функции кэпа и связанных с ним белков:
· участие в сплайсинге;
· участие в процессинге 3'-конца мРНК;
· экспорт мРНК из ядра;
· защита 5'-конца транскрипта от экзонуклеаз;
· участие в инициации трансляции.
Полиаденилирование
Фермент поли(А)-полимераза присоединяет 3'-концу транскрипта от 100 до 200 остатков адениловой кислоты. Полиаденилирование осуществляется при наличии сигнальной последовательности 5'- AAUAAA-3' на 3'-конце транскрипта, за которой следует 5'-CA-3'. Вторая последовательность является сайтом разрезания.
Сплайсинг
После полиаденилирования мРНК подвергается сплайсингу, в ходе которого удаляются интроны (участки, которые не кодируют белки), а экзоны (участки, кодирующие белки) сшиваются и образуют единую молекул. Сплайсинг катализируется крупным нуклеопротеидным комплексом — сплайсосомой, состоящей из белков и малых ядерных РНК. Многие пре-мРНК могут быть подвергнуты сплайсингу разными путями, при этом образуются разные зрелые мРНК, кодирующие разные последовательности аминокислот (альтернативный сплайсинг).
Метилирование
мРНК эукариот подвергаются посттранскрипционному метилированию. То есть сметению (метилированию) наружу ингибиторного гена. Наиболее распространённой модификацией является метилирование остатков аденина по положению N6 с образованием N6-метиладенозина (m6A). Этот процесс метилируют ферменты N6-аденозинметилтрансферазы, которые распознают остатки аденина в консенсусных последовательностях GAC (70 % случаев) и AAC (30 % случаев). Соответствующие деметилазы ингибируют обратный процесс деметилирования. Учитывая обратимость и динамичность процесса метилирования мРНК, а также повышенную концентрацию m6A в длинных экзонах и вокруг стоп-кодонов, предполагают, что метилирование мРНК выполняет регуляторную функцию.
Дата: 2019-05-28, просмотров: 415.