Скотт Менакер
В предыдущих главах обсуждались функциональная анатомия, механика и биофизика дыхательной системы, управляющие обменом О2 и СО2 между вдыхаемым воздухом и циркулирующей кровью. Чтобы потребление О2 и продукция СО2 соответствовали разнообразным требованиям организма, связанным с повседневной активностью, а величины Ра()2 и РаС()2сохранялись в узких физиологических пределах, необходимы приспособительные изменения минутной вентиляции (Vl<;). Для достижения этого гомеостатического эффекта существует сложная система регуляции дыхания.
Эта глава посвящена главным образом хемо- и механорецепторам системы регуляции дыхания. Кроме того, рассматриваются ее интегрированные ответы на значительные патофизиологические возмущения.
Обзор системы регуляции дыхания
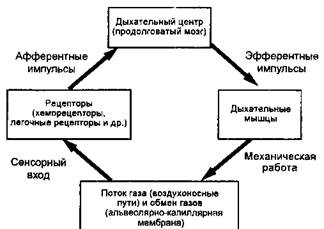
Подобно многим физиологическим системам контроля, система управления дыханием организована как контур отрицательной обратной связи (рис. 16-1). Вдыхаемый газ поступает по ВП к альвеолам, где он участвует в обмене газов на уровне альвеолярно-капиллярной мембраны (гл. 9). Рецепторы откликаются на информацию о механических явлениях (например, о наполнении легких) и гуморальных параметрах (например, Ра()2 и РаС()2). Эта информация интегрируется в дыхательном центре продолговатого мозга, который модулирует нервный импульс к мотонейро-мам, иннервирующим дыхательные мышцы. Координированное возбуждение респираторных мотонейронов приводит к синхронному сокращению дыхательных мышц, создающему воздушный поток.
Когда химические возмущения, подобные гипоксии или гиперкапнии, распознаются хеморецепторами, их сигналы в дыхательном центре реализуются повышенной нервной импульсацией к респираторным мотонейронам, что вызывает повышение минутной вентиляции. Артериальная гипокапния, напротив, вызывает уменьшение вентиляции.
Гуморальная регуляция дыхания
Дыхательная система поддерживает адекватные уровни О2 в артериальной крови, обеспечивая метаболизм периферических тканей, и предотвращает накопление СО2, количество которого увеличивается в процессе метаболизма. Высокоспециализированные ткани контролируют уровень О2 в артериальной крови и уровень СО2 в обильно перфузируемом органе — головном мозге. Эти два сенсорных механизма обеспечивают быстрое изменение характера вентиляции в зависимости от отклонений в газообмене.
Рис. 16-1. Система регуляции .мхамим как копту р отри uaKvib-ioii обратной свя:ш
Центральные хеморецепторы
Из нескольких хемосенсоров, контролирующих обмен газов, ответственность за обнаружение отклонений в гомеостазисе СО2 лежит в основном на центральных хеморецепторах. Несмотря на то, что нейроны, частота импульсации и возбудимость которых модулируются отклонениями Рсо2, обнаружены во многих областях центральной нервной системы, клетки, способные передавать информацию в дыхательный центр, расположены в продолговатом мозге.
Центральные хеморецепторы располагаются вблизи вентральной поверхности продолговатого мозга, где корешки подъязычного нерва выходят из мозгового ствола. Хотя их точное расположение, нервные контакты и нейрохимия изучены недостаточно, эти хемочувствительные клетки, вероятно, отличаются от соседних нейронов, которые обеспечивают дыхательный ритмогенез и составляют основу дыхательного центра.
Хемочувствительные клетки реагируют на отклонения в [1-Г ] и Рсо2 во внеклеточной жидкости внутримозгового интерстициального пространства. Ранее измерения в цереброспинальной жидкости использовались для оценки химизма внутри-мозгового интерстициального пространства. Последние исследования с применением рН-чувствительных электродов показали, что увеличение минутной вентиляции тесно связано с повышением [Н+] во внеклеточной, а не в спинномозговой жидкости.
Когда местная [H^J увеличивается, что эквивалентно падению рН, дыхательный центр увеличивает вентиляцию по сигналу от центральных хеморецепторов. Однако отклонения во внеклеточной [ tT j - не единственный стимул для центральных хеморецепторов. Рост вентиляции в ответ на изолированное повышение внеклеточной [Н+] в отсутствие изменений Рсо-, (изокапнический метаболический ацидоз) происходит медленнее. Кроме того, вентиляторная реакция меньше по величине (рис. 16-2), чем наблюдаемая при изолированном повышении Рсо^ (дыхательный ацидоз).
Одной из причин высокой скорости вентиляторного ответа на гииеркапнию является легкость диффузии СО2 через барьерную систему кровь-головной мозг. Однако этот барьер относительно непроницаем для ионов II* и | НСО < ]. Более того, повышенное РС()-г вызывает расширение сосудов, особенно церебральных, способствуя тем самым усилению диффузии СО2 через гемато-энцефалический барьер. Диффузия СО2 представляет собой еще один важный механизм усиления связи отклонений в кислотно-основном состоянии организма с центральными хеморецеп-торами.
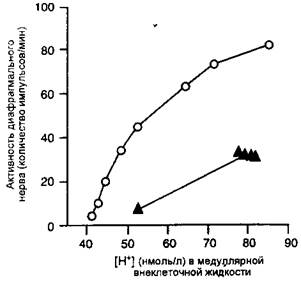
Рис. 16-2. Отношение между |11*| но и не клеточной жидкости продолговатого мо.чга и частотой импульсации или "минутной активностью" диаф-рагмалыюго нериа. |1Г| в медуллярной внеклеточной жидкости повышалась при дыхательном ацидозе (кружки) и метаболическом ацидозе путем ипфузии HCI (треугольники). Вентиляторная реакция на гинеркампию сильнее, чем на метаболический ацидоз, несмотря на сходство отклонений в |1Г| медуллярной внеклеточной жидкости. (Из: lildridge I;.L., KileyJ.P., Millhorn D.H. Respiratory responses to medullary hydrogen ion changes in cats: Different effects of respiratory and metabolic acidosis. J. Physiol. 358: 285-297, 1985.)
При дыхательном ацидозе повышенное РаС( )2 вызывает увеличение диффузии СО2 через барьер кровь-головной мозг. Повышенное СО2 приводит к росту [Н+] около центральных хеморецепторов, которые обнаруживают это изменение и сигнализируют в дыхательный центр о повышении вентиляции, компенсирующем дыхательный ацидоз. Поскольку вентиляторный ответ на увеличение [Н+], обусловленное повышением РС()2, больше, чем ответ на идентичное отклонение [ЬГ] в отсутствие роста Рсо, (рис, 16-2), то РС()2 и [Н+] должны представлять независимые стимулы центральных хеморецепторов. Молекулярные механизмы рецепции остаются неизвестными: либо [Н*] и РС()2 образуют различные стимулы для единого сенсорного механизма, либо специфические стимулы для различных сенсорных механиз-
мов.
Поскольку закисление мозговой интерстициальной жидкости является важным этапом центральной хеморецепции, необходимо выделить главные факторы, влияющие на этот процесс.
В отличие от крови, церебральная интерстициальная жидкость содержит очень мало белка. Без белков с их буферной способностью сдвиги рН во внутримозговой внеклеточной жидкости происходят быстрее, чем в крови. Кроме того, компенсаторный вентиляторный ответ на хроническое закисление мозговой внеклеточной жидкости (как при хронической гиперкапнии) наступает быстрее, чем при закисле-нии крови. Перемещение [HCCV] из крови через гемато-энцефалический барьер в мозговую интерстициальную жидкость происходит влечение 24—48 ч. Задержка [ НСО;< ] в почках в ответ на закисление плазмы занимает от 48 до 72 ч.
Карогидные тельца
В то время как мониторинг РС()2 происходит в стволе головного мозга, отслеживание Р( )2 является функцией исключительно каротидныхтелец (рис. 16-3). Они расположены у бифуркации общих сонных артерий на внутреннюю и наружную (рис. 16-3 А). Несмотря на свой миниатюрный размер, каротидные тельца получают огромный кровоток - от 1.4 до 2.0 л/мин на 100 г ткани, что соответствует особой роли этого органа, как главного датчика, чувствительного к колебаниям кислорода в артериальной крови.
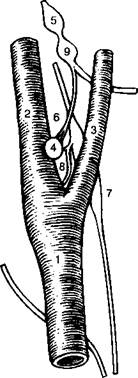
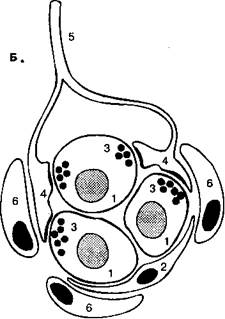
Рис. 16-3. Каротидиос тельце. (А) бифуркация правой сонной артерии (вид спереди). Общая сонная артерия (1) делится па наружную (2) и внутреннюю (.4). Каротидное тельце (4) иппсрвирустся чувствительными волокнами, исходящими из каменистого ганглия (5) и проходящими в составе нерва каротидпого синуса (ft). Верхний шейный ганглий (7) тоже интернирует каротидное тельце посредством гапглиогломсрулярных нервов (8). Узловатый ганглий (9) расположен снаружи внутренней сонной артерии. (Б) Каротидиое тельце, состоит из хеморецепторпых (1) и поддерживающих (2) клеток. В цитоплазме хсморецепторпых клеток находятся сииаптические пузырьки (Л), которые тесно соприкасаются с чувствительными окончаниями (4) ветвящихся волокон нерва каротидпого синуса (5). Вся совокупность этих клеток окружена капиллярами (ft). (Из: (ion/ale/. С'., Alinarax L, Obeso Д., Ritual R. Oxygen and acid chemorcccption in the carotid body chemoreceptors. Trends in Neuroscicncc 15: I/I ft-153/1992.)
Дата: 2019-03-05, просмотров: 342.