| Символ | Трактование символа | ||||
| D u , D | weake | , D | el | Группа слабовыраженных антигенов D | |
| D w | Антиген D Wiel, D-подобный антиген, встречающийся | ||||
| у негров; не следует путать с Dweak | |||||
| D II, D IV, D IVa, | |||||
| DFR, DBT, D HMi | Частичные (парциальные) D-антигены | ||||
| и др. | |||||
| D IV (C) − | Редкий фенотип с парциальным антигеном D IV, слабовыраженным ан- | ||||
| тигеном С и отсутствием антигена е | |||||
| Редкий антиген, встречающийся у лиц с фенотипом D IV (C) − со слабо- | |||||
| R Har | выраженным парциальным антигеном D, реагирующим лишь с неко- | ||||
| торыми сыворотками анти-D, слабовыраженным антигеном C и отсут- | |||||
| o | |||||
| ствием антигена е | |||||
| f (ce, hr) | Антиген f, встречающийся у лиц, унаследовавших гены c и e на одной | ||||
| или обеих гомологичных хромосомах в позиции цис | |||||
| Ce (rhi) | Антиген rh , встречающийся у лиц, имеющих комбинацию антигенов | ||||
| Ce (ген С иi е в позиции цис). | |||||
| D**, *D* | Фенотип с отсутствием антигенов C, E, c, e | ||||
| и сильновыраженным антигеном D | |||||
| D − −, −D − | Фенотип с отсутствием антигенов C, E, c, e (подобно D**), отличаю- | ||||
| щийся от D** наличием редкого антигена Tar (RH40) | |||||
|
| |||||
| R N | Редкий фенотип CDe, при котором антигены Rh, включая D, ослаблены | ||||
| (C)D(e) | Редкий фенотип CDе (подобный R N), при котором антигены Rh осла- | ||||
| блены, а D, наоборот, усилен | |||||
|
| |||||
| – – –, Rhnull | Отсутствие антигенов Rh-Hr (нулевой фенотип) | ||||
157
Система резус полиморфна. Помимо четко очерченных антигенов, она вклю-чает варианты, при которых антигены выражены слабо либо вовсе не продуци-руются. Для ясности дальнейшего изложения объясним некоторых обозначе-ния, встречающиеся в современных публикациях.
Как видно из табл. 4.5, наименования отдельным вариантам, в том числе редко встречающимся , присваивали в значительной мере произвольно. В этом плане классификация ISBT внесла определенный порядок. Тем не менее обо-значения, характеризующие необычную выраженность антигенов или их не-ожиданное отсутствие, в литературе сохраняются, например фенотипы Rhnull, −D −, (C)D(e). В последнем случае необычные фенотипы со слабовыраженны-ми антигенами С и е, кодируемые геном R N и чаще встречающиеся у негров, обозначают как (С)D(e), выделяя скобками очень слабые или практически от-сутствующие антигены С и е.
Обозначение f (ce) и rhi (Ce) с дублирующим синонимом, помещенным в скобки, более информативно для читателя, чем обозначение этих антигенов как f и rhi, поскольку указывает на генетическую подоплеку их формирования (по-зицию цис генов ce или Ce). Антиген f продуцируется комбинацией генов с и е в положении цис. При размещении генов с и е в позиции транс антиген f не фор-мируется. Аналогичная ситуация имеет место в отношении антигена rhi, кото-рый вырабатывается в том случае, если как минимум на одной из унаследован-ных гомологичных хромосом в позиции цис расположены локусы С и е. Гены С и е в позиции транс антигена rhi (Ce) не производят.
Антигены резус встречаются с частотой: D – 85 %, С – 70 %, с – 80 %, E – 30 %, е – 97,5 %. В табл. 4.6 представлены варианты фенотипов и генотипов Rh,
О также результаты серологических реакций, в которые вступают эритроциты с тем или иным сочетанием антигенов резус. Фенотип Rh-Hr выявляют с помощью 5 сывороток: анти-D, анти-C, анти-E, анти-c и анти-e. Сыворотки анти-се, анти-Се, анти-сЕ и анти-СЕ обнаруживают на эритроцитах дополнительный антиген-ный продукт, кодируемый генами, когда они находятся в одном гаплотипе одно-временно. Реагирование этих сывороток при одинаковом фенотипе, но разном ге-нотипе людей не совпадает, что может быть использовано для установления ге-нотипа Rh по фенотипу. Например, лица с фенотипом СсDEe (Ce +ce −cE + CE −),
с большой степенью вероятности (99,99 %) имеют генотип СDe / cDE (генотипы Сde / cDE или СDe / cdE менее вероятны), а лица с тем же фенотипом СсDEe (но Ce −ce +cE −CE + ) имеют генотип СDE / cde или, что менее вероятно, СdE / cDe.
Выраженность антигенов Rh на эритроцитах варьирует в широком диапазо-не. Выделяют сильные, средние и слабые формы антигенов. Эритроциты, не-сущие эти формы, обычно не имеют качественных различий, но отличают-ся от образца к образцу степенью агглютинабельности. Выраженность агглю-тинации (агглютинабельность) определяется количеством антигена, представ-ленного на поверхности эритроцитов, что обусловлено генетическими фактора-ми. Агглютинабельность эритроцитов людей с генотипом cDE / cDE выражена
158
сильнее, чем эритроцитов лиц с генотипом CDe / CDe, поскольку количество ан-тигенных участков на эритроцитах DE больше, чем на эритроцитах DC. Редкий фенотип −D −, при котором отсутствуют антигены С, Е, с и е, отличается наибо-лее высоким содержанием субстанции D по сравнению с нормальным D-типом. Менее всего антиген D выражен на эритроцитах со слабым D-фенотипом (D u) и совсем не выражен на эритроцитах Rhnull.
В редких случаях варианты агглютинабельности могут быть обусловлены качественными различиями парциальных антигенов, которые содержат непол-ный набор D-эпитопов.
Таблица 4.6
Фенотипы и генотипы Rh
Реакция ( +, −) с антителами | Частота, | Частота, | ||||||||||||
| к антигену | Фенотип | Генотип | ||||||||||||
| % | % | |||||||||||||
| D | C | E | c | e | ce | Ce | cE | CE | C W | |||||
| CDe / cde | 29,90 | |||||||||||||
| + | + | − | + | + | + | + | − | − | − | CcDe | 31,93 | CDe / cDe | 1,98 | |
| cDe / Cde | 0,05 | |||||||||||||
| + | + | − | − | + | − | + | − | − | − | CDe | 16,81 | CDe / Cde | 16,01 | |
| CDe / Cde | 0,80 | |||||||||||||
| + | + | − | + | + | − | − | CDe / cDE | 12,24 | ||||||
| + | + + | + + | + + | + | + | − | + | + | − | CDE / cDe | 0,01 | |||
| + | + | − | + | + | − | − | CDe / cdE | 0,97 | ||||||
| + | + + | + + | + + | + | − | + | + | − | − | CcDEe | 13,69 | cDE / Cde | 0,27 | |
| + | + + | + + | + + | + | + | − | − | + | − | CDE / cde | 0,19 | |||
| + | − | − | − | + | + | − | cDE / СdE | 0,006 | ||||||
| cDE / cde | 10,04 | |||||||||||||
| + | − | + | + | + | + | − | + | − | − | cDEe | 11,82 | cDE / cDe | 0,72 | |
| cDe / cdE | 0,06 | |||||||||||||
| + | + | + | − | + | − | + | − | + | − | CDEe | 0,07 | CDE / Cde | 0,07 | |
| CDE / Cde | ||||||||||||||
| + | − | + | + | + | − | − | + | − | − | cDE | 2,49 | cDE / cDE | 2,49 | |
| cDE / cdE | ||||||||||||||
| + | − | − | + | + | + | − | − | − | − | cDe | 2,21 | cDe / cde | 2,21 | |
| cDe / cDe | ||||||||||||||
| + | + | + | + | − | − | − | + | + | − | CcDE | 0,035 | CDE / cdE | 0,035 | |
| cDE / CdE | ||||||||||||||
| + | + | + | − | − | − | − | − | + | − | CDE | 0,00 | CDE / CDE | 0,00 | |
| + | + + | + + | − | + | − | + | + | − | + | C WCDEe | C W DE / Cde | |||
| + | − | + | − | − | − | + | + | C W Dе / CDЕ | ||||||
| C W De / CDe | ||||||||||||||
| + | + | − | − | + | − | + | − | − | + | C WCDe | C W De / Cde | |||
| C W de / CDe | ||||||||||||||
| + | − | − | + | + | + | − | − | − | + | C WcDe | 2 – 9 | C W De / cDe | 2–9 | |
| C W De / cde | ||||||||||||||
| + | − | − | + | + | + | − | − | − | + | C W DE / cde | ||||
| + | − | − | + | + | + | − | − | − | + | C WcDEe | C W dE / cDe | |||
| + | − | − | + | + | − | − | − | − | + | C W De / cDE | ||||
| + | − | − | − | + | − | − | − | − | + | C WC WDe | C W De / C W De | |||
| C W De / C W de | ||||||||||||||
Резус-
принадлежность
| Ре зу с–п ол ож и т е л ь н ы е |
159
Окончание табл. 4.6
| Реакция ( +, −) с антителами | Частота, | Частота, | Резус- | |||||||||||||
| к антигену | Фенотип | Генотип | ||||||||||||||
| % | % | принадлежность | ||||||||||||||
| D | C | E | c | e | ce | Ce | cE | CE | C W | |||||||
| − | − | − | + | + | + | − | − | − | − | cdе | 13,5 | cde / cde | 13,5 | |||
| − | + | − | + | + | + | − | − | − | − | Ccdе | 2,14 | Cde / cde | 2,14 | е | ||
| − | + | − | − | + | + | − | − | − | − | Cde | Cde / Cde | |||||
| ы | ||||||||||||||||
| − | − | + | + | + | + | − | + | − | − | cdEe | 0,27 | cdE / cde | 0,27 | ь н | ||
| − | − | − | + | − | − | − | + | − | − | cdE | cdE / cdE | е л | ||||
| цат | ||||||||||||||||
| − | + + | + + | + | + | − | + | + | − | − | CcdEe | Cde / cdE | |||||
| − | + | + | + | + | − | + | − | CdE / cde | и | |||||||
| − | + | + | − | − | − | − | − | + | − | CdE | 0,08 | CdE / CdE | 0,08 | р | ||
| –от | ||||||||||||||||
| − | + | + | + | − | − | − | + | − | − | CcdE | СdE / cdE | |||||
| с | ||||||||||||||||
| − | + | + | − | + | − | + | − | + | − | CdEe | CdE / Cdе | |||||
| Ре зу | ||||||||||||||||
| − | + | + | − | + | − | − | − | + | + | C WCdEe | 0,00 | C W de / CdE | ||||
| − | − | − | + | + | + | − | − | − | + | C Wcde | 0,00 | C W de / cde | 0,00 | |||
| − | − | − | − | + | − | − | − | − | + | C Wde | 0,00 | C W de / C W de | ||||
Генетика
Система Rh – одна из наиболее полиморфных систем, антигены которой ко-дируются 2 генами (RHD и RHCE), расположенными на коротком плече хро-мосомы 1 в локусе RH между 1p34.3 и 1p36.13 (Cherif-Zahar и соавт. [211], MacGeoch и соавт. [453], Marsh и соавт. [462]).
Три генетические теории
Существуют 3 генетические концепции наследственной передачи антигенов Rh. Первая разработана в начале 50-х годов прошлого столетия Александром Винером (Wiener и соавт. [714, 715]), вторая – в тот же период Рональдом Фишером совместно с Робертом Рейсом (Fisher, Race [283, 284], Race [543]). Третья концепция, получившая в последние годы подтверждение, предложена в 90-х годах прошлого столетия Патрисией Типпетт (Tippett [654, 656]).
Интересно проследить логику построения этих концепций.
Располагая двумя сыворотками: анти-Rhо и анти-rh', выявляющими 2 антиге-на – Rhо и rh', Винер вполне обоснованно допустил, что существует не 2, а 4 аг-глютиногена резус: Rhо, rh', Rhо' и rh-агглютиноген, не содержащий ни Rhо, ни rh'. Он полагал, что аллель R о гена Rh обусловливает продукцию антигена Rhо, аллель R 1 – продукцию антигенов Rhо и rh', аллель r' – антигена rh', а аллель r – отсутствие обоих антигенов – Rhо и rh'.
Винер, не имея экспериментальных данных, свидетельствующих о том, что отдельные гены могут быть сцеплены, сделал вывод , что все антигенные признаки Rh контролирует только 1 (но полиморфный) ген (рис. 4.1). Это и явилось основой его концепции, получившей известность как теория одно-го гена.
160

Рис. 4.1. Генетические концепции системы резус.
1 – теория одного гена A. Wiener (1943),
2 – теория трех генов R. Fisher и R. Race 1946),
3 – теория двух генов P. Tippett (1986).
Согласно представлениям Винера, разнообразие факторов резус и их сочетаний в наблюдаемых фенотипах людей обусловлено одним геном, который встречается в виде 8 аллеломорфных вариантов: R 1, R 2, R о, R Z, r, r', r" (см. табл. 4.4).
Обнаружение новых факторов (E, e, C W и др.), легко укладывались в концеп-цию Винера. Так, после открытия антигена hr' (c) и hr" (e) он дополнил первона-чальный постулат, указав, что ген R о может кодировать помимо Rhо еще и фак-торы hr' и hr" (фенотип Rоr, или cDe). Открытие антигена C W Винер трактовал как обнаружение нового аллеля R 1w, а открытие антигена hr S, присутствующего у большинства людей hr" (e), – как новый аллель R^h о.
Однако по мере открытия новых антигенов и их разновидностей обозначе-ния Винера стали затруднительными.
середине 50-х годов иммуносерологи располагали уже не двумя, а пятью сы-воротками антирезус, дифференцирующими соответствующие антигены резус: D, C, E, c и е. С помощью этих сывороток американской школой исследователей во главе с Винером и английской школой во главе с Фишером и Рейсом были установ-лены 8 гаплотипов, кодирующих различные сочетания антигенов (см. табл. 4.4).
В отличие от Винера, утверждавшего, что ген резус неделим, Фишер и Рейс полагали, что существуют 3 сцепленных локуса (теория трех генов), которые наследуются одновременно. Согласно их представлению, в этих локусах на хро-мосоме в линейном порядке расположены гены D, C и E, кодирующие антигены D, C и E (см. рис. 4.1). Доссе [50] добавил четвертую пару аллельных генов Ff.
В этих же локусах располагаются соответствующие им 3 аллельных гена: d, c и е, кодирующие антитетичные антигены d, c и е. В каждом локусе может присут-ствовать один ген: D или d, C или c, E или e. Таким образом, каждый индивид получает с хромосомой матери и отца от 3 до 5 антигенных признаков, опреде-ляющих его резус-фенотип.
161
Фишер сформулировал понятие об антитетичных антигенах и предсказал существование антигенов Е, е и d. Предвидение двух первых вскоре (в 1943 г.) блестяще подтвердили Wiener, Sonn [713], открыв антиген Е, и Mourant [494] (в 1945 г.) – е. Третий гипотетический антиген (d) так и не был обнаружен. Антитела анти-d были описаны Hill и Haberman в 1948 г. [349], Matthes в 1950 г. [467], однако эти находки не подтвердились. По-видимому, указанные авторы исследовали неизвестные в то время антитела анти-се, реагирующие с эритро-цитами cde, но не реагирующие с эритроцитами CDe, cDE, что создавало ви-димость анти-d-специфичности. Как сегодня известно (см. далее), у лиц Rh −
с локусе, симметричном D, генетический материал в виде аллеля d не найден. Отсутствие гена d объясняет, почему многочисленные целенаправленные пои-ски антигена d не увенчались успехом.
Теория трех генов позволила установить последовательность генов в генном локусе RH, а также объяснить происхождение редких фенотипов резус.
В соответствии с этой теорией локусы RH располагаются в последователь-ности Dd – Cc – Ee (см. рис. 4.1). По мнению Фишера и Рейса, разрыв генно-го комплекса происходит на дистанции С – E чаще, чем на дистанции D – C, из чего следует, что гены D и E наиболее удалены друг от друга. Доказательством такого расположения генов явилась находка фенотипа −D − (Race и соавт. [552], Read и соавт. [556]), для которого характерно отсутствие антигенов Сс и Ее. Если бы ген D располагался между генами C и E, Cc – Dd – Ee, то делеция ло-куса Cс и Eе в варианте −D − должна была так или иначе сказаться на гене D. Однако, напротив, антиген D в фенотипе −D − серологически более выражен, чем в фенотипе CDe и cDE, что связано, по-видимому, с отсутствием конку-рентного влияния генов С и Е на D вследствие их делеции.
Согласно теории трех генов, редкие генные комбинации (dCe, dcE, DCE, dCE) возникают в результате кроссинговера частых генных комбинаций: DCe, DcE, dce. На рис. 4.2 приведены варианты кроссинговера. Если крос-синговер происходит между хромосомами, несущими гaплотипы DСe и dce, образуется редкий гаплотип dCe; если между хромосомами, несущими га-плотипы DcE и dce, – редкий гаплотип dcE; если между DСe и DcE – DСE. При повторном кроссинговере DCE и dce образуется еще более редкий га-плотип – dCE. Как видно из рис. 4.2, гаплотип Dce образуется при каждом из перечисленных вариантов кроссинговера. Сумма частоты (2,49 %) ред-ко встречающихся в европеоидных популяциях фенотипов (dCe, dcE, DCE, dCE) и частота фенотипа Dce (2,6–3 %) примерно совпадали, что подтверж-дало правильность теории в целом.
Относительная редкость кроссинговера (≈ 3 %), по мнению Фишера и Рейса, свидетельствует о том, что гены DCE близко расположены друг к другу и крос-синговер между ними происходит скорее как исключение, чем как правило.
Большинство исследователей нашли CDE-терминологию более удобной для повседневного использования, чем Rh-Hr, и она получила статус прикладной.
162
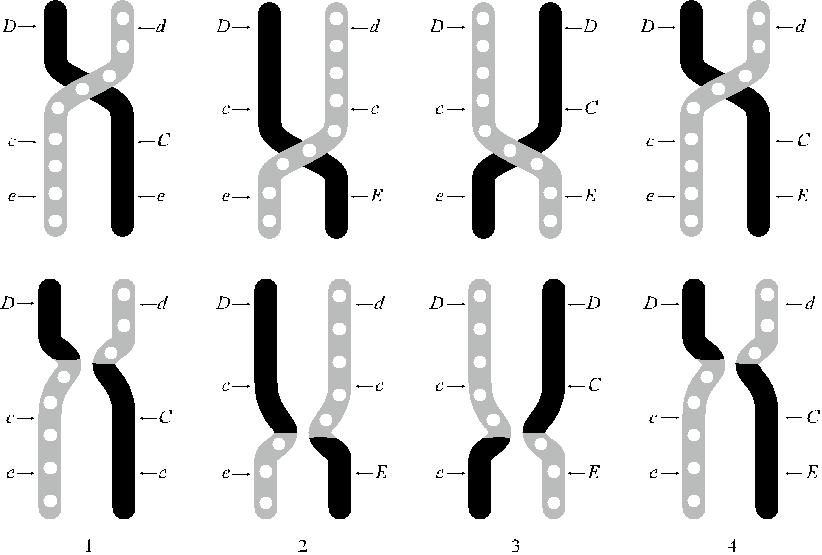
Рис. 4.2. Варианты кроссинговера по Фишеру и Рейсу.
1 и 4 – перекрест в участке D – C, 2 и 3 – перекрест в участке С – Е.
с результате кроссинговера возникают следующие гаплотипы: 1 – dCe и Dce, 2 – dcE и Dce, 3 – DCE и Dce, 4 – dCE и Dce.
Благодаря своей простоте теория трех генов Фишера – Рейса завоевала всеобщее признание и несколько потеснила концепцию Винера, которая от-нюдь не утратила своего значения и практического применения до настояще-го времени.
Спустя годы концепцию трех тесно связанных локусов модифицировали в однолокусную, которая предполагает 3 сублокуса в одном генном комплексе.
Открытие Sanger и соавт. [596] антигенов f (ce) и rhi (Ce) выходило за рам-ки концепции трех генов, постулирующей принцип «один ген – один антиген». Следовало признать, что система Rh включает не 3, а 4 (C, D, E + ce), затем 5 (C, D, E, ce + Ce) локусов. Перекрестно реагирующие сыворотки и необычные Rh-фенотипы также не вписывались в теорию трех генов и кроссинговера по Фишеру. Номенклатура Фишера – Рейса стала затруднительной для обозначе-ния большого числа новых Rh-антигенов и ей на помощь пришла цифровая но-менклатура, предложенная Розенфельдом.
Свои концепции Винер, Фишер и Рейс основывали исключительно на ре-зультатах серологических исследований (популяционные и семейные). Концепция Типпетт (теория двух генов) также основана на данных серологиче-ских исследований, однако более широкого масштаба. К началу 90-х годов арсе-нал сывороток, которым располагали иммуносерологи, составил более 40 наи-менований, включая анти-се, анти-Се, анти-сЕ и анти-СЕ. Накопились много-численные данные о качественном и количественном разнообразии фенотипов Rh, в том числе данные о биохимической природе Rh-антигенов.
Moore и соавт. [482] в 1982 г. и Ridgwell и соавт. [564] в 1983 г. независи-мо друг от друга нашли, что Rh-антигены располагаются на двух протеинах
163
мембраны эритроцитов: один белок несет на себе антиген D, другой – C и E. Имеющиеся сведения послужили толчком к формированию новой, современ-ной молекулярно-генетической модели системы Rh, предложенной Типпетт.
Согласно концепции Типпетт [654, 656], система Rh контролируется дву-мя тесно сцепленными структурными генами , один из которых кодирует D-антиген, другой – антигены С, с, Е и е. Первый ген включает 2 аллеля: D, по-лучивший название RHD, и не-D (отсутствие кодирующего субстрата); второй ген представлен четырьмя аллелями: се, Се, сЕ и СЕ. Продукты второго гена, получившего название RHCE, идентифицируют с помощью 5 специфических сывороток: анти-D, анти-се, анти-Се, анти-сЕ и анти-СЕ (табл. 4.7). По мнению Типпетт, отдельные эпитопы полипептида СсЕе (С, с, Е и е) более иммуноген-ны, чем их комплексы (се, Се, сЕ и СЕ ), в связи с чем антитела анти-с , анти-С, анти-Е и анти-е встречаются чаще, чем комплексные антитела анти-се, анти-Се, анти-сЕ, анти-СЕ.
Данные, представленные Типпетт (см. табл. 4.7), с такой же убедительно-стью свидетельствуют о существовании четырех аллелей гена СЕ, как и данные, полученные в свое время Wiener, Fisher и Race с помощью пяти специфических сывороток: анти-D, анти-с, анти-С, анти-Е и анти-е. Вопрос о том, какая кон-цепция ближе к действительности, сегодня решается в пользу взглядов Типпетт, подтверждающихся данными молекулярно-биологических исследований.
Необычные, в том числе редкие, фенотипы резус, как полагает Типпетт, воз-никают в результате мутаций, делеций и транслокаций генетического веще-ства. На это указывают фенотипы с ослабленными антигенами (С)(е) или (с)(е) (табл. 4.8), которые ассоциированы с редкими антигенами – Rh32, Rh35, Rh36, Rh48. Мутации и другие воздействия на генный локус RH нарушают продук-цию нормального антигена, создают новые необычные формы антигенов.
Таблица 4.7
Реакции генного продукта основных 8 гаплотипов*
| Генный | Реакция с сыворотками | ||||
| продукт | анти-D | анти-ce | анти-Ce | анти-cE | анти-CE |
| Dce | + | + | – | – | – |
| DCe | + | – | + | – | – |
| DcE | + | – | – | + | – |
| DCE | + | – | – | – | + |
| dce | + | + | – | – | – |
| dCe | – | – | + | – | – |
| dcE | – | – | – | + | – |
| dCE | – | – | – | – | + |
* По Tippett [654].
164
Таблица 4.8
Дата: 2019-02-24, просмотров: 581.