1. Общая физиология синапса.
2. Локализация медиаторов и соответствующих нейронов в ЦНС.
3. Свойства химических синапсов.
Общая физиология синапса
Термины «синапс» и «синаптическая передача» были введены в физиологию Ч. Шеррингтоном в 1897 году. Исследуя деятельность ЦНС, он предположил, что между собой нейроны сообщаются с помощью специального синаптического механизма. Синапс – это морфофункциональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (рис. 3). Все синапсы ЦНС можно классифицировать следующим образом.
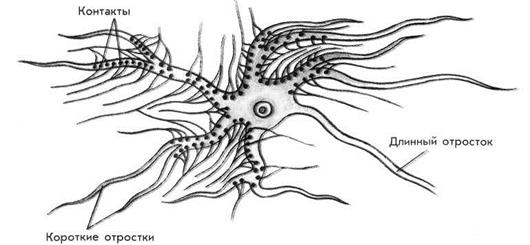
Рис. 3. Расположение контактов на нейроне.
1. По локализации – центральные (головной и спинной мозг) и периферические (нервно-мышечный, нейро-секреторный, синапс вегетативной нервной системы). Центральные синапсы можно, в свою очередь, разделить на: аксо-аксональные, аксо-дендритические (дендритные), аксо-соматические, дендро-дендритические, дендро-соматические и т.п.
2. По развитию в онтогенезе – стабильные (например, синапсы дуг безусловного рефлекса) и динамичные, появляющиеся в процессе индивидуального развития.
3. По конечному эффекту – тормозные (тормозящие) и возбуждающие.
4. По механизму передачи сигнала – электрические, химические, смешанные.
Химические синапсы можно классифицировать по:
а) форме контакта – терминальные (колбообразные соединения) и проходящие (варикозные расширения аксона);
б) природе медиатора – холинергические (медиатор – ацетилхолин), адренергические (норадреналин, в отдельных случаях адреналин), дофаминергические (дофамин), серотонинергические (серотонин), ГАМК-ергические (медиатор – гамма-аминомасляная кислота), глицинергические (глицин), глютаматергические (глютамат), пептидергические (медиатор – пептиды), пуринергические (медиатор – АТФ), азотергические (медиатор – оксид азота NО) и другие.
Электрические синапсы (эфапсы). В основном они локализованы в стволе мозга, в том числе они обнаружены между нейронами мезенцефального ядра тройничного нерва, а также между нейронами вестибулярного ядра Дейтерса и ядер нижней оливы в продолговатом мозге. С точки зрения морфологии электрический синапс представляет собой щелевидное образование с ионными мостиками-каналами между двумя контактирующими клетками. Петли тока, при наличии потенциала действия (ПД), почти беспрепятственно проходят через такой щелевидный контакт и возбуждают, т.е. индуцируют, генерацию ПД в соседней клетке. В целом, синапсы обеспечивают очень быструю передачу возбуждения. Но в то же время с помощью этих синапсов нельзя обеспечить одностороннее проведение, т.к. большая часть таких синапсов обладает двусторонней проводимостью. Кроме того, с их помощью нельзя вызывать торможение эффекторной клетки.
Аналогом электрического синапса в гладких мышцах и в сердечной мышце являются щелевые контакты типа нексуса.
Химические синапсы представляют собой окончания аксона (терминальные синапсы) или его варикозную часть (проходящие синапсы).
Химический синапс состоит из трех компонентов (или элементов): пресинаптической части, постсинаптической части и синаптической щели. В пресинаптической части содержится медиатор, который под влиянием нервного импульса выделяется в синаптическую щель и, связываясь с рецепторами в постсинаптической части, вызывает ряд физиологических эффектов, в том числе изменение ионной проницаемости постсинаптической мембраны, что приводит к ее деполяризации (в возбуждающих синапсах) или гиперполяризации (в тормозных синапсах).
Пресинаптическая часть представляет собой расширенную конечную часть аксона. В ней содержатся митохондрии, агранулярная эндоплазматическая сеть, нейрофиламенты, нейротрубочки и синаптические пузырьки диаметром 20–65 нм, в которых находится нейромедиатор.
Постсинаптическая часть представлена постсинаптической мембраной, содержащей синаптические рецепторы (мембранные рецепторы), связывающиеся с нейромедиатором. Мембрана утолщена за счет скопления под ней плотного филаментозного белкового материала.
Синаптическая щель – ее ширина варьирует от 20–30 нм до 50 нм.
Любой химический синапс, независимо от природы медиатора и хеморецептора, активируется под влиянием потенциала действия, распространяющегося к пресинапсу от тела нейрона. Под влиянием потенциала действия происходит деполяризация пресинаптической мембраны, что повышает проницаемость кальциевых каналов пресинаптической мембраны и приводит к увеличению входа в пресинапс ионов Са2+. В ответ на это происходит высвобождение (выход из пресинапса) 100–200 порций (квантов) медиатора, что осуществляется путем экзоцитоза. Выйдя в синаптическую щель, медиатор взаимодействует со специфическими рецепторами постсинаптической мембраны. Активированные медиатором рецепторы непосредственно регулируют проницаемость ионных каналов постсинаптической мембраны. Взаимодействие медиатора с постсинаптическими рецепторами изменяет ионную проницаемость. В синапсах, в которых осуществляется возбуждение постсинаптической структуры, обычно происходит повышение проницаемости для ионов Na+ или Са2+, что вызывает деполяризацию постсинаптической мембраны. Эта деполяризация получила название возбуждающего постсинаптического потенциала (или ВПСП). Если его величина достигает критического уровня деполяризации, то во внесинаптических областях генерируется ПД. В тормозных синапсах в результате взаимодействия медиатора с рецепторами, наоборот, происходит гиперполяризация (за счет, например, увеличения проницаемости для ионов калия и хлора). Этот вид изменения мембранного потенциала получил название тормозного постсинаптического потенциала (или ТПСП). В гиперполяризованном состоянии клетка снижает свою возбудимость и благодаря этому прекращает отвечать на внешние раздражители или уменьшает спонтанную активность.
Одновременно выделившийся в синаптическую щель медиатор может взаимодействовать с рецепторами, расположенными на пресинаптической мембране. Таким способом регулируется интенсивность последующего высвобождения медиатора, т.е. процесс экзоцитоза. Это получило название антидромного эффекта (или явления обратной связи).
После каждого цикла проведения нервного импульса медиатор разрушается с участием специфического фермента и подвергается удалению. Одновременно происходит обратный захват медиатора (например, норадреналина) или продуктов его расщепления в пресинаптическую либо в постсинаптическую структуру.
Синтез медиатора совершается в пресинаптическом элементе, куда из крови или спинно-мозговой жидкости попадают исходные продукты (предшественники медиаторов) и ферменты, необходимые для его синтеза. Ферменты образуются в соме нейрона и по аксону, примерно со скоростью 6 мм/сутки, транспортируются в пресинаптическое окончание аксона, где используются в процессе синтеза медиатора. Угнетение активности этих ферментов фармакологическим путем может привести к истощению запасов медиатора в синапсе и снижению его функциональной способности. Затем образовавшийся медиатор путем активного транспорта вводится в синаптические везикулы (мелкие везикулы).
Синтез нейропептидов происходит подобно синтезу пептидных гормонов. Первоначально крупные аминокислотные последовательности образуются на рибосомах и помещаются в эндоплазматический ретикулюм. В цистернах аппарата Гольджи осуществляется протеолитический процесс расщепления крупных полипептидов на фрагменты с образованием активных пептидов, которые включаются в отпочковавшиеся крупные везикулы. Разные пептидные фрагменты могут оказаться в различных везикулах, которые транспортируются в нервные окончания нейрона. Синтез и упаковка в везикулы классических медиаторов и нейропептидов в нейроне происходят параллельно. Поэтому из его нервных окончаний освобождаются несколько различных медиаторов.
В основе освобождения медиатора из синапса лежит процесс экзоцитоза, который представляет собой разновидность активного транспорта, предназначенного в живых системах для выделения в окружающую среду гормонов, медиаторов, модуляторов и других веществ.
В роли медиатора выступают десятки и даже сотни химических веществ. Для того чтобы назвать вещество медиатором, используют следующие критерии: 1. Вещество выделяется из клетки при ее активации. 2. В клетке имеются ферменты для синтеза данного вещества. 3. В соседних клетках имеются белки-рецепторы, активируемые данным медиатором. 4. Фармакологический (экзогенный) аналог имитирует действие медиатора. Наряду с медиаторами в синапсе есть модуляторы, т.е. вещества, которые прямо не участвуют в процессе передачи сигнала от нейрона к нейрону, но могут, однако, этот процесс существенно усиливать или ослаблять.
2. Локализация медиаторов и соответствующих
нейронов в ЦНС
В разных отделах ЦНС расположены нейроны, имеющие различные медиаторы.
Ацетилхолин является медиатором альфа-мотонейронов спинного мозга и ствола (их аксоны заканчиваются нервно-мышечным синапсом), нейронов коры больших полушарий, ретикулярной формации мозга, преганглионарных нейронов вегетативной (симпатической и парасимпатической) нервной системы, ганглионарных нейронов парасимпатической нервной системы, а также нейронов сетчатки.
Дофамин является медиатором нейронов, сконцентрированных в среднем мозге (черная субстанция, покрышка мозга), в гипоталамусе, в симпатических ганглиях и в сетчатке. Аксоны дофаминергических нейронов достигают нейронов базальных ганглиев, лимбической системы, коры больших полушарий.
Норадреналин является медиатором нейронов, локализованных в голубом пятне ствола мозга (с проекцией в кору мозга, гипоталамус, мозжечок, спинной мозг), а также ганглионарных нейронов симпатической нервной системы.
Серотонин является медиатором серотонинергических нейронов, локализованных, главным образом, в ядрах шва ствола мозга (проекция аксонов в кору головного мозга, гипоталамус, мозжечок, спинной мозг) и в сетчатке.
Гистамин является медиатором нейронов сосцевидных телец гипоталамуса. Аксоны гистаминергических нейронов проецируются в кору мозга, таламус, базальные ганглии, мозжечок, спинной мозг.
Глютамат является медиатором возбуждающих нейронов, локализованных в различных отделах коры больших полушарий, в таламусе, базальных ганглиях, мозжечке, гипоталамусе, стволе мозга, спинном мозге, а также в сетчатке.
Глицин является медиатором тормозных нейронов, расположенных главным образом в спинном мозге и сетчатке.
ГАМК является медиатором тормозных нейронов коры мозга, мозжечка, ствола мозга, спинного мозга (совместно с глицином) и сетчатки.
АТФ является медиатором возбуждающих нейронов уздечки головного мозга, спинного мозга, афферентных нейронов, а также симпатических нейронов.
В последние годы в ЦНС выявлено большое число нейроактивных пептидов. Так же, как и медиаторы (нейротрансмиттеры), пептиды идентифицируются как нейроактивные вещества. Для них характерна следующая локализация в ЦНС. Субстанция Р – головной мозг, окончания афферентных нейронов в ноцицептивной системе. Вазопрессин, окситоцин – задний гипофиз, продолговатый мозг, спинной мозг. Кортиколиберин – медиальное возвышение гипоталамуса и другие отделы мозга. Тиреолиберин – гипоталамус, сетчатка. Соматолиберин – гипоталамус. Соматостатин – гипоталамус и другие отделы мозга, желатинозная субстанция, сетчатка. Эндорфины – гипоталамус, таламус, ствол мозга, сетчатка. Нейротензин – гипоталамус, сетчатка. Гастрин – гипоталамус, продолговатый мозг. Глюкагон – гипоталамус, сетчатка. Мотилин – нейрогипофиз, кора мозга, мозжечок. Секретин – гипоталамус, таламус, обонятельная луковица, ствол мозга, кора мозга, перегородка, гиппокамп, стриатум.
Принцип Дейла. Английский нейрофизиолог, работавший в 1930–1950-е годы, Дж. Дейл сформулировал принцип «Один нейрон – один медиатор». Он полагал, что каждый нейрон выделяет один и тот же медиатор из всех своих (даже далеко удаленных) нервных окончаний. Доказательством тому служили данные о том, что медиатор ацетилхолин выделяется как из окончаний длинного аксона, который альфа-мотонейрон посылает к скелетной мышце, так и из короткого ответвления, которое, не покидая спинного мозга, активирует клетку Реншоу. Однако уже в 1960–1980-е годы было показано, что выделение многих классических медиаторов (ацетилхолина, норадреналина, ГАМК) сопровождается одновременным высвобождением веществ – комедиаторов (нейропептидов). Например, в парасимпатических окончаниях одновременно с ацетилхолином выделяется вазоактивный интестинальный пептид. При этом медиатор обычно оказывает быстрое действие, а комедиатор – медленное или даже только модулирующее. Поэтому было предложено правило Дейла видоизменить: «Один нейрон – один быстрый медиатор».
Анализ тормозной передачи в спинном мозге вскоре заставил пересмотреть и это положение. Оказалось, что в большинстве тормозных синапсов спинного мозга одновременно из одного нейрона выделяются две быстрые тормозные аминокислоты – ГАМК и глицин. Новая редакция принципа Дейла стала звучать так: «Один нейрон – один быстрый синаптический эффект». В 1999 году были получены данные о том, что в спинном мозге из одного нейрона может выделяться быстрый тормозной медиатор (ГАМК) и быстрый возбуждающий медиатор (АТФ). Таким образом, можно полагать, что число и вид медиаторов, а также комедиаторов и модуляторов в синапсе может быть различным.
Нарушение продукции медиатора – достаточно распространенное явление среди людей. При этом основные симптомы, наблюдаемые в условиях дефицита медиатора, определяются его функцией. Так, при нарушении синтеза ацетилхолина развивается миастения и старческая деменция (болезнь Альцгеймера). Нарушение продукции дофамина вызывает развитие болезни Паркинсона и шизофрении. При нарушении продукции норадреналина и серотонина наблюдаются депрессии, галлюцинации, нарушения сна. Снижение выработки гистамина вызывает вегетативные нарушения, а глютамата – эпилепсию, моторные нарушения, нарушения памяти, дегенеративные нарушения; глицина – судорожный синдром; ГАМК – хорею, судорожный синдром, депрессию; АТФ – нарушение болевой чувствительности, сосудистые расстройства; аденозина – судорожные состояния.
Дата: 2019-02-02, просмотров: 1316.