В развитии нервной системы многоклеточных принято выделять три этапа (или три типа) нервной системы – диффузную (кишечно-полостные), узловую (членистоногие) и трубчатую (позвоночные).
В процессе эволюции структуры и функции нервной системы происходили совершенствование, с одной стороны, ее отдельных элементов, в том числе нервных клеток, а с другой – ее общих физиологических свойств. Все это вместе обеспечивало расширение адаптационных возможностей организма. Рассмотрим особенности каждого из трех выделенных этапов эволюции нервной системы.
Диффузная, или сетевидная, нервная система. Первым этапом развития нервной системы было формирование нервной системы диффузного типа, нервные клетки которой мало напоминают нейроны позвоночных. В них отсутствует субстанция Ниссля, ядро не дифференцировано, количество отростков невелико, их длина незначительна. Передача возбуждения от нейрона к нейрону осуществляется не только с помощью синапсов, но и через протоплазматические мостики. Нейроны слабо дифференцированы по функции. Скорость распространения возбуждения по нервным волокнам предельно низкая и составляет сотые и десятые доли метра в секунду.
Таким образом, основными особенностями нервной системы диффузного типа являются неопределенность связей, отсутствие четко выраженных входов и выходов, надежность функционирования. Для этого этапа эволюции нервной системы характерен и низкий коэффициент использования энергии.
Узловая нервная система как второй этап в эволюции этой системы. Нейроны узлового типа нервной системы коренным образом отличаются от элементов диффузного типа. На этом этапе филогенеза произошли количественные и качественные изменения нейронов – возросло их общее число и увеличилось число их разновидностей, возникло большое количество вариаций нейронов, отличающихся по размеру, форме и числу отростков. Формирование нервных узлов привело к обособлению и структурной дифференцировке трех основных видов нейронов – афферентных (чувствительных), ассоциативных (вставочных) и эфферентных (двигательных).
В пределах нервной системы узлового типа (у насекомых, крабов, дождевого червя) имеются также нервные волокна, покрытые многослойной оболочкой, которая напоминает миелиновую оболочку у нервных волокон позвоночных. Скорость проведения в таких волокнах намного выше, чем в аксонах такого же диаметра беспозвоночных, но меньше, чем у миелинизированных аксонов большинства позвоночных. Перехваты Ранвье здесь короче, чем у позвоночных. Именно этим обусловлена меньшая скорость распространения возбуждения по сравнению с аналогичным волокном у позвоночных. Как известно, механизм сальтаторного (скачкообразного) проведения возбуждения – довольно позднее эволюционное приобретение. Такой способ проведения возбуждения характерен лишь для позвоночных животных, имеющих нервную систему трубчатого типа.
Трубчатая нервная система. Нервная система трубчатого типа – высший этап структурной и функциональной эволюции нервной системы. Все позвоночные, начиная от самых примитивных форм (ланцетник) и заканчивая человеком, имеют центральную нервную систему в виде нервной трубки, оканчивающейся в головном конце головным мозгом. На уровне млекопитающих развивается кортикализация – процесс формирования новой коры, экранных структур, обладающих свойствами, которые существенно отличают ЦНС млекопитающих от ЦНС других животных.
ЦНС позвоночных состоит из спинного и головного мозга. Трубчатый вид имеет только спинной мозг. Головной мозг, развиваясь как передний отдел нервной трубки и проходя стадии мозговых пузырей, к моменту созревания претерпевает значительные конфигурационные изменения при существенном нарастании объема.
Спинной мозг при своей морфологической непрерывности в значительной степени сохраняет принцип сегментарности и метамерности брюшной нервной цепочки узловой нервной системы. Так, простые сухожильные рефлексы сохраняются в случае целостности только 2–3 сегментов спинного мозга, замыкающих эту рефлекторную дугу. С другой стороны, чрезвычайно развитый аппарат внутри спинномозговых связей обеспечивает целостность и богатство спинно-мозговых рефлексов. Среди них имеются сложные цепные рефлексы (например, шагательный), вовлекающие в процесс возбуждения практически весь спинной мозг. В процессе эволюции позвоночных животных вместе с прогрессирующим усложнением структуры и функции спинного мозга нарастает его зависимость от головного. Это хорошо видно на примере явления спинального шока. Оно проявляется в торможении деятельности спинного мозга после отделения его от головного мозга. Такое торможение деятельности спинного мозга обусловлено в первую очередь прекращением нисходящих корригирующих влияний со стороны головного мозга. Чем сильнее эти влияния, тем более глубоки и длительны последствия их прекращения. Например, у лягушки явление спинального шока прекращается очень быстро – уже спустя несколько минут, у кошки время восстановления функций спинного мозга исчисляется часами, у приматов – неделями и месяцами. Травматический перерыв спинного мозга у человека приводит к развитию тяжелейших, практически необратимых проявлений спинального шока.
Процесс энцефализации, т.е. совершенствования структуры и функций головного мозга, у млекопитающих дополняется кортикализацией – формированием коры больших полушарий и совершенствованием ее функций. Если на уровне стволовых отделов и базальных ганглиев переднего мозга нейроны сгруппированы в виде отдельных специализированных ганглиев или ядер, то кора дает примеры совершенно новых принципов структурной и функциональной организации. Построенная по экранному принципу кора больших полушарий содержит не только специфические проекционные, но и значительные по площади ассоциативные зоны. Последние служат для корреляции различных сенсорных влияний, их интеграции с прошлым опытом для того, чтобы по моторным путям передать сформированные паттерны (рисунки) возбуждения и торможения для реализации поведенческих актов.
В отличие от ганглионарных структур кора головного мозга обладает рядом свойств, характерных только для нее. Важнейшее из них – чрезвычайно высокая пластичность и надежность, как структурная, так и функциональная. Изучение этих свойств центральной нервной системы в эволюции позвоночных позволило А.Б. Когану в 60-х годах XX века обосновать вероятностно-статистический принцип организации высших функций мозга. Этот принцип в наиболее яркой форме выступает в коре головного мозга, являясь одним из приобретений прогрессивной эволюции.
Онтогенез нервной системы
Нервная система плода начинает развиваться на ранних этапах эмбриональной жизни. Из наружного зародышевого листка – эктодермы – по спинной поверхности туловища эмбриона образуется утолщение – нервная трубка. Головной конец этой трубки развивается в головной мозг, остальная часть – в спинной мозг.
У недельного эмбриона намечается незначительное утолщение в оральном (ротовом) отделе нервной трубки. На третьей неделе зародышевого развития в головном отделе нервной трубки образуются три первичных мозговых пузыря (передний, средний и задний), из которых развиваются главные отделы головного мозга – конечный, средний, ромбовидный.
В последующем передний и задний мозговые пузыри расчленяются каждый на два отдела, в результате чего у четырех-пяти-недельного эмбриона образуется пять мозговых пузырей: 1) конечный (телэнцефалон), 2) промежуточный (диэнцефалон), 3) средний (мезэнцефалон), 4) задний (метэнцефалон), 5) продолговатый (миелэнцефалон) (рис. 1). В последующем из конечного мозгового пузыря развиваются полушария головного мозга и подкорковые ядра; из промежуточного пузыря – межуточный мозг (зрительные бугры, подбугорье); из среднего пузыря формируется средний мозг – четверохолмие, ножки мозга, сильвиев водопровод; из заднего пузыря – мост мозга (варолиев мост) и мозжечок; из продолговатого пузыря – продолговатый мозг. Задняя часть миелэнцефалона плавно переходит в спинной мозг.
Из полостей мозговых пузырей и нервной трубки образуются желудочки головного мозга и канал спинного мозга. Полости заднего
и продолговатого мозговых пузырей превращаются в IV желудочек, полость среднего мозгового пузыря превращается в узкий канал – водопровод мозга (сильвиев водопровод), который сообщает между собой III и IV желудочки. Полость промежуточного пузыря превращается в III желудочек, а полость конечного пузыря – в два боковых желудочка. Через посредство парного межжелудочкового отверстия III желудочек сообщается с каждым боковым желудочком; IV желудочек сообщается с каналом спинного мозга. В желудочках и спинно-мозговом канале циркулирует спинно-мозговая жидкость.

 Нейроны развивающейся нервной системы посредством своих отростков устанавливают связи между различными отделами головного и спинного мозга, а также входят в связь с другими органами. Чувствительные нейроны, входя в связь с другими органами, заканчиваются рецепторами – периферическими окончаниями, воспринимающими раздражение; двигательные нейроны заканчиваются синапсом – контактным образованием нервного волокна с мышцей.
Нейроны развивающейся нервной системы посредством своих отростков устанавливают связи между различными отделами головного и спинного мозга, а также входят в связь с другими органами. Чувствительные нейроны, входя в связь с другими органами, заканчиваются рецепторами – периферическими окончаниями, воспринимающими раздражение; двигательные нейроны заканчиваются синапсом – контактным образованием нервного волокна с мышцей.
К третьему месяцу внутриутробного развития выделяются основные части центральной нервной системы: большие полушария и ствол мозга, мозговые желудочки, а также спинной мозг. К пятому месяцу дифференцируются основные борозды коры больших полушарий, однако кора остается еще недостаточно развитой. На шестом месяце отчетливо выявляется функциональное превалирование высших отделов нервной системы плода над нижележащими отделами.
Головной мозг новорожденного имеет относительно большую величину, и масса его в среднем составляет 1/8 массы тела, т.е. около 400 г, причем у мальчиков она несколько больше, чем у девочек.
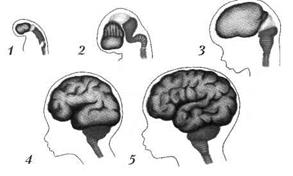
У новорожденного хорошо выражены борозды, крупные извилины, однако их глубина и высота невелики. Мелких борозд относительно мало, они появляются постепенно в течение первых лет жизни. К девяти месяцам первоначальная масса мозга удваивается и к концу первого года составляет 1/11–1/12 массы тела. К трем годам масса головного мозга по сравнению с массой его при рождении утраивается, к пяти годам она составляет 1/13–1/14 массы тела. К двадцати годам первоначальная масса мозга увеличивается в четыре-пять раз и составляет у взрослого человека всего 1/40 массы тела. Рост мозга происходит в основном за счет миелинизации нервных проводников и увеличения размера имеющихся уже при рождении примерно
20 млрд нервных клеток. Наряду с ростом головного мозга меняются пропорции черепа (рис. 1).
Мозговая ткань новорожденного малодифференцирована. Корковые клетки, подкорковые узлы, пирамидные пути недоразвиты, плохо дифференцируются на серое и белое вещество. Нервные клетки плода и новорожденного расположены концентрированно на поверхности больших полушарий и в белом веществе мозга. С увеличением поверхности головного мозга нервные клетки мигрируют в серое вещество; концентрация их от общего объема мозга уменьшается. В то же время плотность мозговых сосудов увеличивается.
У новорожденного затылочная доля коры больших полушарий имеет относительно большие размеры, чем у взрослого. Количество полушарных извилин, их форма, топографическое положение претерпевают определенные изменения по мере роста ребенка. Наибольшие изменения происходят в течение первых пяти-шести лет и заканчиваются лишь к пятнадцати-шестнадцати годам. Боковые желудочки мозга сравнительно широкие. Соединяющее оба полушария мозолистое тело тонкое и короткое. В течение первых пяти лет оно становится толще и длиннее. К двадцати годам мозолистое тело достигает окончательных размеров.
Мозжечок у новорожденного развит слабо, расположен высоко, имеет продолговатую форму, малую толщину и неглубокие борозды. Варолиев мост постепенно, по мере роста, перемещается к скату затылочной кости. Продолговатый мозг новорожденного расположен более горизонтально. Черепно-мозговые нервы расположены симметрично на основании мозга.
В послеродовом периоде претерпевает изменения и спинной мозг. По сравнению с головным спинной мозг новорожденного имеет более законченное морфологическое строение, в связи с чем он оказывается более совершенным в функциональном отношении.
Спинной мозг новорожденного относительно длиннее, чем у взрослого. В дальнейшем рост спинного мозга отстает от роста позвоночника, в связи с чем его нижний конец «перемещается» кверху. Рост спинного мозга продолжается приблизительно до двадцати лет. Его масса увеличивается за это время примерно в восемь раз.
Окончательное соотношение спинного мозга и позвоночного канала устанавливается к пяти-шести годам. Рост спинного мозга наиболее выражен в грудном отделе. Шейное и поясничное утолщения спинного мозга начинают формироваться в первые годы жизни ребенка. В этих утолщениях сконцентрированы клетки, иннервирующие верхние и нижние конечности. С возрастом отмечается увеличение количества клеток в сером веществе спинного мозга, наблюдается и изменение их микроструктуры. Спинной мозг имеет густую сеть венозных сплетений, что объясняется относительно быстрым ростом вен спинного мозга по сравнению с темпами его роста.
Периферическая нервная система новорожденного недостаточно миелинизирована, пучки нервных волокон редкие, распределены неравномерно. Процессы миелинизации происходят неравномерно в различных отделах. Миелинизация черепно-мозговых нервов наиболее активно осуществляется в течение первых трех-четырех месяцев и заканчивается к году. Миелинизация спинно-мозговых нервов продолжается до двух-трех лет. Вегетативная нервная система функционирует с момента рождения. В дальнейшем отмечаются слияние отдельных узлов и образование мощных сплетений симпатической нервной системы.
На ранних этапах эмбриогенеза между различными отделами нервной системы формируются четко дифференцированные, «жесткие» связи, создающие основу для жизненно необходимых врожденных реакций. Набор этих реакций обеспечивает первичную адаптацию после рождения (например, пищевые, дыхательные, защитные реакции). Взаимодействие нейронных групп, обеспечивающих ту или иную реакцию либо комплекс реакций, составляет функциональную систему.
3. Возрастная эволюция мозга
и ее принципы гетерохронности
В процессе онтогенетического развития мозг человека претерпевает значительные изменения. В анатомическом отношении мозг новорожденного и мозг взрослого человека существенно отличаются. Это значит, что в процессе индивидуального развития происходит возрастное эволюционирование мозговых структур. Даже после завершения морфологического созревания нервной системы человека остается необъятная «зона роста» в смысле совершенствования, перестройки и нового образования функциональных систем. Мозг как совокупность нервных элементов остается у всех людей примерно одинаковым; однако на основе этой первичной структуры создается бесконечное разнообразие функциональных особенностей.
В процессе эволюции мозга выделяют два важнейших стратегических направления. Первое из них заключается в максимальной предуготованности организма к будущим условиям существования. Это направление характеризуется большим набором врожденных, инстинктивных реакций, которыми организм оснащен буквально на все случаи его жизни. Но набор таких «случаев» довольно стереотипен и ограничен: питание, защита, размножение.
В рамках второго направления эволюции происходит неуклонное увеличение размеров коры больших полушарий мозга. Этот отдел является наименее специализированным и наиболее пригодным
для фиксации личного опыта. Принцип кортикализации функций предполагает возможность их непрерывного совершенствования.
На каждом возрастном этапе какие-то функции или отдельные звенья нервно-психического развития выглядят наиболее активными и сформированными. Наступает следующий возрастной период – и картина меняется: недавние «лидеры» отходят на вторые места, появляются новые формы и способы реагирования.
Например, новорожденный ребенок обладает набором первичных автоматизмов, обеспечивающих прежде всего акт сосания и регуляцию мышечного тонуса. Зрительное, слуховое восприятия находятся еще в рудиментарном состоянии. Но постепенно зрительные реакции становятся все более активными: от автоматической фиксации взгляда на случайно попавшем в поле зрения предмете ребенок переходит к самостоятельному зрительному поиску; он приобретает способность разглядывать предмет, «ощупывать» его взглядом. К шестому-седьмому месяцу жизни разглядывание становится важнейшим способом изучения окружающего мира. Однако вскоре, как только появляется возможность брать предметы, перекладывать их из одной руки в другую (девять-десять месяцев), активное манипулирование приобретает главную роль в деятельности ребенка. С появлением речи мануальное (ручное) познание все более вытесняется словесным.
Если какой-то возрастной этап представить как финишную черту, то можно увидеть, что к данному финишу различные функциональные системы приходят с разной степенью зрелости, совершенства. Одни уже почти оформились и в дальнейшем лишь незначительно модифицируются, другие только начинают формироваться. В этом заключается принцип гетерохронности, неодновременности созревания отдельных функциональных систем мозга. Зрительное восприятие, например, совершенствуется быстрее, чем слуховое или вкусовое, а способность понимать обращенную речь возникает гораздо раньше, чем умение говорить.
Гетерохронность развития отдельных звеньев функциональной системы регистрируют при помощи анатомо-физиологических исследований. Тем самым объективно раскрывается материальный субстрат процессов развития мозга. В частности, большое внимание уделяется темпам миелинизации периферических нервов – скорости образования миелиновой оболочки в нервных проводниках. Миелиновая оболочка является эволюционным приобретением, позволяющим проводить нервные импульсы с большей скоростью и более дифференцированно. Миелинизированные нервные волокна обнаруживаются только у представителей относительно поздних этапов эволюции и в наибольшей степени – у млекопитающих, включая человека. Сопоставление степени миелинизации у взрослых и детей различных возрастов показывает, сколь неравномерно происходит этот процесс в различных отделах нервной системы. Например, волокна лицевого нерва, участвующие в обеспечении акта сосания, оказываются миелинизированными уже к моменту рождения, а пирамидный путь, связывающий двигательные центры коры головного мозга с соответствующими отделами спинного мозга, завершает миелинизацию лишь к двум годам. Процессы миелинизации косвенно отражаются на скоростях проведения импульсов по волокнам нерва. Эти скорости определяются при помощи электронейромиографии.
Установлено, что общая тенденция, характерная для созревания нервной системы, заключается в увеличении скоростей проведения нервных импульсов. Темпы прироста скоростей в разных отделах нервной системы неодинаковы в различные возрастные периоды. Так, у новорожденных наиболее высокие скорости проведения обнаруживаются в тех волокнах лицевого нерва, которые связаны с актом сосания. Эти показатели даже мало отличаются от величин, характерных для взрослого человека. Скорости проведения в нервах верхних и нижних конечностей новорожденного значительно ниже. В дальнейшем отмечается быстрое нарастание скоростей проведения импульсов в верхних конечностях, что предшествует появлению у ребенка манипулятивной деятельности. К восьми-девятимесячному возрасту, когда возникают попытки самостоятельно вставать на ноги, обнаруживается бурный прирост скоростей проведения в нижних конечностях. Этот прирост опережает соответствующие показатели для верхних конечностей вплоть до того периода, пока ребенок не овладеет самостоятельной ходьбой. В дальнейшем скорости проведения импульсов в верхних конечностях снова начинают расти быстрее и раньше достигают характерных для взрослых норм.
Из всех этих данных следует, что гетерохрония нарастания скоростей проведения отчетливо связана с усложнением двигательных функций. Схема «лицо–руки–ноги–руки» соответствует основным этапам моторного развития ребенка. Кроме того, нарастание скоростей проведения предшествует формированию новой функции. В этом проявляется принцип опережающего обеспечения функции, характерный для развивающейся нервной системы.
4. Методы исследования функций ЦНС
и функциональных состояний мозга
Функции центральной нервной системы изучают с использованием классических для общей физиологии и специальных методов.
Метод раздражения. Заключается в том, что на определенные структуры ЦНС наносят раздражение электрическим током или химическими веществами. Электрическая стимуляция мозга осуществляется через введенные в мозг электроды в «острых» опытах на животных или во время хирургических операций на мозге у человека. У человека электрическая стимуляция мозга применяется для изучения связи между психическими процессами и функциями и отделами мозга. Так, например, можно изучать физиологические основы речи, памяти, эмоций.
Метод раздражения различных участков ЦНС электрическим током обычно выполняется с использованием стереотаксической техники. Она предусматривает введение электродов в мозг человека или животного в строго определенные его участки и на определенную глубину через отверстия, просверленные в черепе.
В лабораторных и клинических условиях используется метод микрополяризации, т.е. пропускание слабого постоянного тока через отдельные участки коры головного мозга. При этом электроды прикладываются к поверхности черепа в области стимуляции. Локальная микрополяризация не разрушает ткань мозга, а лишь оказывает влияние на сдвиги потенциала коры в стимулируемом участке, поэтому она может быть использована в психофизиологических исследованиях. Клиническим вариантом этого метода является электронаркоз, широко применяемый в акушерстве для лечения слабости родовой деятельности.
При изучении механизма действия различных химических веществ на нейроны мозга, в том числе наркотиков в опытах на животных, используется введение этих веществ через специальную канюлю в соответствующие участки мозга.
Методы удаления (экстирпации) и разрушения (повреждения) определенных участков центральной нервной системы для установления их функций в обеспечении поведения – один из наиболее старых и распространенных методов изучения физиологических основ поведения. В ряде случаев эти методы применяют в клинических условиях с лечебной целью, т.е. по медицинским показаниям. С их помощью можно удалить очаг, вызывающий эпилептические припадки. Пионером в этом направлении был У. Пенфильд. В России этот метод нашел применение в клинике академика Н.П. Бехтеревой при лечении ряда форм патологии ЦНС, в том числе при болезни Паркинсона. Конечно, использование этого метода для лечения человека имеет целый ряд ограничений. В последние годы с лечебными целями в клинике проводится разделение полушарий путем рассечения межполушарной связки, т.е. мозолистого тела (коллозиотомия).
Методы регистрации электрической активности структур мозга широко используются при исследовании функций ЦНС и ее высших отделов – коры больших полушарий. В этом аспекте используют разнообразные методы, в том числе методы внутриклеточного и внеклеточного отведения электрической активности отдельных нейронов, а также методы внеклеточного отведения суммарной электрической активности мозга (например, метод энцефалографии, или ЭЭГ).
Методы исследования рефлекторной деятельности спинного и головного мозга представляют собой наиболее важные методы физиологических наблюдений и клинических исследований. В зависимости от целей исследований изучают проявление различных безусловных рефлексов (например, при изучении общих свойств ЦНС) или осуществляют выработку условных рефлексов при исследовании работы коры больших полушарий. Так, изучая на лягушках защитный сгибательный рефлекс, возникающий при погружении лапки животного в раствор кислоты, И.М. Сеченов открыл наличие процессов торможения в ЦНС, а А.А. Ухтомский, исследуя рефлекторную деятельность двигательной коры у кошек, открыл явление доминанты – одного из важнейших свойств ЦНС.
Морфологические (включая гистологические, гисто- и цитохимические, электронно-микроскопические), биохимические, биофизические, радиоизотопные, радиоиммунологические и другие методы исследования широко используются при изучении физиологии ЦНС, а также ее отдельных систем – вегетативных, сенсорных, двигательных и интеллектуальных.
Методы оценки состояния вегетативной нервной системы. Существует много методик, используемых в физиологических исследованиях и в клинической практике с целью оценки состояния вегетативной нервной системы (ВНС). Большая часть из них основана на исследовании проявления различных вегетативных рефлексов. Например, при оценке кожных рефлексов исследуют пиломоторный рефлекс, или рефлекс «гусиной кожи» (вызывается болевым или холодовым раздражением кожи в области трапециевидной мышцы), и потовые рефлексы (аспириновая проба, пилокарпиновая проба, проба Минора, исследование кожно-гальванического рефлекса). При оценке сосудистых рефлексов часто исследуется местный дермографизм (за счет штрихового раздражения кожи предплечья или других частей тела) и болевой дермографизм. При оценке зрачковых рефлексов исследуют прямую и содружественную реакцию зрачков на свет, реакции зрачков при конвергенции (или схождение глазных яблок), при аккомодации, т.е. при рассматривании предмета вблизи, реакцию на боль. При оценке висцеральных рефлексов исследуют глазо-сердечный рефлекс Данини–Ашнера (изменение сердечного ритма при надавливании на глазные яблоки), клиностатический и ортостатический рефлекс Превеля (изменение сердечного ритма, вызываемое переходом человека соответственно из вертикального положения в горизонтальное, или из горизонтального в вертикальное), рефлекс Геринга (изменение сердечного ритма при произвольной задержке дыхания).
Методы оценки состояния двигательных систем мозга.
В физиологических наблюдениях и в клинической практике для оценки состояния двигательных систем мозга исследуют двигательные функции. Для этих целей у человека оцениваются объем и сила произвольных движений, мышечный тонус, проявление нормальных безусловных двигательных рефлексов (корнеальный, конъюнктивальный, глоточный, небный, брюшные рефлексы, сухожильные рефлексы, в том числе бицепс-рефлекс, трицепс-рефлекс, коленный рефлекс, ахиллов рефлекс), а также наличие патологических безусловных рефлексов (рефлексы Бабинского, Оппенгейма, Гордона, Шеффера, Россолимо, Бехтерева, Жуковского и др.), наличие синкинезий (содружественные движения, возникающие в парализованных конечностях) и клонусов (длительные сокращения мышц), проявление защитных рефлексов. Кроме того, оценивают функциональное состояние двигательных ядер черепно-мозговых нервов – глазо-двигательного, блокового, тройничного, отводящего, лицевого, языко-глоточного, блуждающего, добавочного и подъязычного, а также вестибулярных ядер, красного ядра, ретикулярной формации ствола мозга, мозжечка, базальных ганглиев и двигательной коры больших полушарий. В частности, для оценки состояния мозжечка и связанных с ним стволовых образований исследуют способность к поддержанию равновесия и выполнения сложнокоординированных движений, для чего применяют различные пробы, в том числе пробу Ромберга, пальценосовую пробу, пяточно-коленную пробу, диадохокинез, а для оценки состояния двигательной коры больших полушарий проверяют наличие явлений апраксии (моторной, конструктивной, идеомоторной). Для оценки состояния двигательных систем мозга также применяют различные электрофизиологические и миографические методы, включая электромиографию и ЭЭГ.
Методы оценки состояния сенсорных систем мозга. В физиологических наблюдениях и в клинической практике для оценки состояния сенсорных систем мозга применяются различные объективные и субъективные методы исследования. Так, при исследовании зрительной сенсорной системы используются методы оценки рефракции и ее нарушений, субъективные методы определения остроты зрения (по таблицам Д.А. Сивцева, состоящим из буквенных оптотипов или колец Ландольта), объективные методы (скиаскопический метод, метод регистрации оптокинетического нистагма), оценка наличия астигматизма, определение аккомодационной способности хрусталика. Широко используются методы оценки полей зрения и наличия скотом, исследование цветового зрения, оценка глазного дна, оценка бинокулярного зрения и скрытого косоглазия, оценка внутриглазного давления (например, с помощью тонометра Маклакова).
Для оценки состояния слухового анализатора используются методы речевой, тональной (с помощью аудиометров) и камертональной (проба Ринне и проба Вебера с использованием камертонов) аудиометрии. Проводится оценка бинаурального слуха. Состояние вестибулярного анализатора оценивается с помощью таких функциональных проб, как вращательная проба (например, в кресле Барани), проба Ромберга, проба на походку, проба на нистагм. При исследовании состояния тактильной сенсорной системы оценивается простая тактильная чувствительность, в том числе чувство прикосновения, давления и вибрации, а также сложная тактильная чувствительность, в том числе чувство локализации, дискриминационная чувствительность, двумерно-пространственное чувство и трехмерно-пространственное чувство (стереогнозис). Для оценки состояния болевой и температурной сенсорных систем изучают соответственно болевую и температурную чувствительность различных участков кожи путем нанесения болевых и температурных раздражений. Для оценки состояния проприоцептивной сенсорной системы исследуют мышечно-суставную чувствительность, или глубокую чувствительность, субъективными и объективными (например, с использованием кинематометра Жуковского) методами. Для оценки состояния вкусового анализатора определяют вкусовую чувствительность, в том числе по абсолютным порогам чувствительности к сладким, горьким, соленым и кислым веществам (на различных участках языка). Для оценки состояния обонятельной сенсорной системы определяют обонятельную чувствительность, в том числе остроту обоняния (с помощью ольфактометра) по минимальному количеству пахучего вещества, вызывающего ощущение запаха. Более детально большинство из вышеуказанных методов рассматриваются при описании соответствующих сенсорных систем.
Методы исследования ВНД. Для оценки состояния ВНД используют метод условных рефлексов, а также разнообразные инструментальные и неинструментальные методики, в том числе электроэнцефалографию (ЭЭГ) в различных ее вариантах, магнитоэнцефалографию, компьютерную томографию, ядерно-магнитно-резонансную томографию мозга, позитронно-эмиссионную трансаксиальную томографию и функционально-магнитно-резонансную томографию, методы регистрации электрической активности кожи, или кожно-гальванической реакции (КГР), методы регистрации работы отдельных систем и органов, методы тестов, корректурных проб и специализированных опросников, метод моделирования. Кроме того, применяется ряд методических приемов, позволяющих охарактеризовать отдельные свойства нервной системы, в том числе такие, как функциональная подвижность, или лабильность.
Метод условных рефлексов предложен И.П. Павловым. Он позволяет исследовать разнообразные аспекты ВНД, в том числе способность к выработке условных рефлексов, включая положительные и отрицательные, т.е. выработку внутреннего торможения. Благодаря методу условных рефлексов были установлены основные закономерности работы первой и второй сигнальных систем, изучены механизмы формирования условных рефлексов, внутреннего торможения, исследованы процессы анализа и синтеза, а также явлений иррадиации и концентрации процессов возбуждения и торможения в коре больших полушарий. Именно с помощью метода условных рефлексов
И.П. Павлову удалось дать характеристику основных свойств нервной системы и тем самым объективизировать выделение основных типов ВНД. В современной нейрофизиологии метод условных рефлексов применяется в комплексе с другими методами при исследовании механизмов обучения, становления и развития адаптивного поведения, способствуя тем самым более полному представлению о протекающих в мозге физиологических процессах. В клинической практике метод условных рефлексов используется в диагностических и лечебных целях.
Электроэнцефалография – это метод регистрации и анализа биоэлектрической активности мозга.
Регулярная электрическая активность мозга может быть зафиксирована уже у плода и прекращается только со смертью. Даже при глубокой коме и наркозе наблюдается особая характерная картина мозговых волн.
Условия регистрации и способы анализа ЭЭГ следующие.
В стационарный комплекс для регистрации ЭЭГ и ряда других физиологических показателей входят звукоизолирующая экранированная камера, оборудованное место для испытуемого, многоканальные усилители, регистрирующая аппаратура (чернилопишущий энцефалограф, многоканальный магнитофон). Обычно используется от восьми до 16 каналов регистрации ЭЭГ от различных участков поверхности черепа одновременно. Анализ ЭЭГ осуществляется как визуально, так и с помощью ЭВМ. В последнем случае необходимо специальное программное обеспечение.
Компьютерная томография мозга (КТМ) проводится с использованием томографа или специализированного нейротомографа. Она позволяет прижизненно получить точные и детальные изображения изменений плотности мозгового вещества человека на основе применения рентгеновского излучения и компьютерной обработки результатов анализа. Компьютерная томография мозга позволяет определить местоположение опухоли, получить представления о распределении регионального мозгового кровотока и интенсивности обмена веществ в различных структурах головного мозга. О высокой разрешающей способности метода говорит тот факт, что определение максимально активизированных участков мозга может осуществляться
с точностью до 1 мм.
Ядерно-магнитно-резонансная (ЯМР), позитронно-эмиссион-ная трансаксиальная и функционально-магнитно-резонансная томографиямозга являются более совершенными вариантами компьютерной томографии мозга, в которых используется эффект ядерного магнитного резонанса (ЯМР-томография), позитронной эмиссионной томографии (ПЭТ) и функционального магнитного резонанса (ФМР). Эти методы относятся к наиболее перспективным способам неинвазивного комплексного изучения структуры, метаболизма и кровотока мозга.
При ЯМР-томографии получение изображения основано на определении в мозговом веществе распределения плотности ядер водорода (протонов) и на регистрации некоторых их характеристик при помощи мощных электромагнитов, расположенных вокруг тела человека.
Преимущество ЯМР-томографии заключается в отсутствии ионизирующего излучения. С его помощью можно получить четкие изображения «срезов» мозга в различных плоскостях.
Позитронно-эмиссионная трансаксиальная томография (ПЭТ-сканирование) сочетает возможности компьютерной томографии мозга и радиоизотопной диагностики, для которой используются ультракороткоживущие позитронизлучающие изотопы, или «метки» (это естественные метаболиты мозга), которые вводятся через дыхательные пути или внутривенно. С помощью ПЭТ измеряют региональный мозговой кровоток и метаболизм глюкозы или кислорода в отдельных участках головного мозга. ПЭТ позволяет осуществлять прижизненное картирование на «срезах» мозга регионального обмена веществ и кровотока.
Метод функционального магнитного резонанса(ФМР) – это вариант совмещения метода ЯМР с измерением мозгового метаболизма при помощи позитронно-эмиссионной томографии. Его применение расширяет возможности прижизненного исследования структурных и функциональных особенностей мозга.
Методы регистрации работы отдельных систем и органов. При изучении функционального состояния мозга иногда применяются методы регистрации деятельности сердечно-сосудистой и дыхательной систем, мышечной системы. Среди них электрокардиография (регистрация электрической активности сердца), пульсометрия, тонометрия (измерение артериального давления), реография (регистрация изменения сопротивления электрическому току как отражение изменения кровотока в соответствующем органе), плетизмография (регистрация изменения объема органа в связи с изменением его кровенаполнения), пневмография (регистрация дыхательных движений и изменения объема грудной клетки), динамометрия (измерение мышечной силы), электромиография (регистрация электрической активности скелетных мышц), эргография (регистрация работы мышц) и многие другие.
Оценка эмоционального состояния с помощью «детектора лжи». Многие из указанных выше показателей широко применяются в специальном приборе, предназначенном для выявления эмоционального состояния человека. Он называется «детектором лжи» и представляет собой полиграф, одновременно регистрирующий комплекс физиологических показателей (КГР, ЭЭГ, плетизмограмму и др.) с целью выявить динамику эмоционального напряжения. С человеком, проходящим обследование на полиграфе, проводят собеседование, в ходе которого наряду с нейтральными задают вопросы, составляющие предмет специальной заинтересованности. По характеру физиологических реакций, сопровождающих ответы на разные вопросы, можно судить об эмоциональной реактивности человека и в какой-то мере о степени его искренности в данной ситуации. Поскольку в большинстве случаев специально не обученный человек не контролирует свои вегетативные реакции, «детектор лжи» дает по некоторым оценкам до 71% случаев обнаружения обмана.
Неинструментальные методы исследования (тестирование с помощью опросников). В физиологии, психологии и психофизиологии в последние годы широкое распространение получили опросники, корректурные тесты и другие варианты неинструментальных методов исследования. К ним относятся личностный опросник Г. Айзенка, используемый с целью определения экстраверсированности и эмоциональной стабильности (нейротизма), что позволяет оценить тип ВНД человека. Для оценки характера человека применяется Миннесотский личностный опросник (MMPI), предложенный в 1967 году Мак Кинли, а также 16-факторный личностный опросник Кеттела. Для оценки эмоционального состояния, уровня тревожности и стресса применяются тест Тейлора, или шкала манифестации тревоги Тейлора, опросник личностной тревожности Ч.Д. Спилбергера–Ю.Л. Ханина. Для оценки самочувствия, активности и настроения – психологический тест «САН» В.А. Доскина и соавторов, а также цветовой тест Люшера.
Для оценки подвижности нервных процессов (возбуждения и торможения), умственной продуктивности, концентрации и распределения внимания применяются разнообразные корректурные тесты, например, тест В.Я. Анфимова, тест Тулуз–Пьерона. Для исследования мышления и интеллектуальных способностей – тест Равена, тест Векслера, тест на определение показателя интеллекта IQ. Для оценки различных характеристик внимания применяются тест Платонова–Шульте (объем и переключение внимания), тест перепутанных линий (устойчивость и концентрация внимания), для оценки памяти – тест Джекобса (объем кратковременной слуховой памяти), тест Бюллера (объем смысловой памяти).
ЛЕКЦИЯ 5
Дата: 2019-02-02, просмотров: 886.