В истории промысла сардины последние 500 лет известны несколько периодов увеличения ее запасов. Ито (1961) выделил четыре периода: 1568-1580, 1690-1720, 1790-1840, 1910-1950, 1972-1992гг.
В период предыдущей вспышки, т.е. в 20-30 годы, ежегодный вылов добывающими странами (Россия, Япония, Корея) достигал 2,8 млн.т. При этом отечественный вылов у берегов Приморья составлял 142 тыс.т. Сокращение запасов к 1938 году привело к снижению уловов по краям нагульной части ареала, у Сахалина и Хоккайдо почти в 10 раз, однако общий вылов по-прежнему оставался на высоком уровне.
В течение 1941-1942 годов сардина почти исчезла у берегов Приморья, о.Хоккайдо, Курильских островов. Резко упали уловы и на юге Японии. В 1951-1961 годах наблюдалось некоторое увеличение уловов за счет кратковременного роста численности япономорских популяций.
Депрессия численности сардины продолжалась около 30 лет (1941 1972 гг.), а с начала 70-х годов началось новое увеличение запасов.
В отличие от 30-х годов, когда доминировало нерестилище только в районе Сатсуан, где выметывалось до 70% икры, в последний период вспышки численности сардины наблюдалось увеличение нерестовых площадей всех четырех популяций сардины. С конца 70-х годов нерестилища вокруг Японии слились воедино, что способствовало воспроизводству и явилось следствием высокой численности вида. Однако, с 1981 года урожайность поколений стала снижаться что послужило показателем прекращения роста численности сардины.
Отечественный промысел сардины в последний период роста ее численности в тихоокеанских водах у побережья Японии и Курильских островов был организован в 1974 году. Сначала он был нестабильным, эпизодическим и велся в основном у побережья о. Хонсю, между 35° и 37°с.ш.. Годовой вылов составлял всего 2,1 тыс.т. В результате увеличения численности сардины в Тихом океане вылов в дальнейшем значительно возрос и достиг 1979году 357 тыс. т. Затем вылов стал снижаться, но не за счет сокращения численности запасов, а из-за перераспределения флота в другие районы промысла (Японское море). С 1983 года уловы в тихоокеанских водах опять увеличились. В Южно-Курильском районе с 1989 по 1992 год b уловах почти отсутствовали сеголетки, что давало возможность с заблаговременностью предсказать о резком снижении численности сардины в последующие годы.
Отсутствием сеголеток в уловах объясняется увеличение средних размеров сардины в1991 году до 20,5 - 21,7 см (в 1987 г. они составляли 18,6 19,7 см). Последние урожайные поколения были в 1986 - 1987 годах.
При общем сокращении запасов еще больше сократился нагульный ареал, основная часть которого оказалась в зоне Японии.
По предварительным данным в.1993 году общий вылов сардины в Тихом океане должен составить 200 тыс. т; из них в зоне Японии 150-160 тыс. т, в зоне Южных Курил 40-50 тыс.т (биомасса около 150 тыс.т).
В Японском море отечественный промысел был начат в 1976 rоду и достиг максимума в 1990 году (530 тыс.т), а затем уловы стали резко снижаться. В 1991 году промысловая обстановка была неустойчивой, промысел базировался на поколениях 1986-1988 годов рождения.
В уловах отсутствовали особи в возрасте 0+, 1+, в результате чего отмечалось увеличение средних размеров рыб.
Биомасса сардины начала сокращаться с 1988 года. По сравнению с 1988 годом в 1994 году биомасса сардины снизилась в 200 раз. Это связано с тем, что поколения 1990-1992 годов были неурожайными, в результате пополнение промыслового стада было слабым, что отразилось на численности и уловах в последующие годы.
В Атлантическом океане из семейства сельдевых Clupeidae встречаются все три рода сардин и илиша.
Сардинелла круглая Sardinella aurita Valenciennes (рис. 3.1.2.) Отличается от других сардин гладкой жаберной крышкой, на заднем конце которой имеется темное пятно. В спинном плавнике обычно 14 мягких лучей, иногда количество лучей варьирует от 12 до 15. В анальном от 12 до 16 мягких лучей, но обычно - 15. В продольном ряду от 45 до 48 чешуек. Зубов нет, а если имеются, то мельчайшие. Длина достигает 30 см. Работы по исследованию роста сардинеллы, проводившиеся Наварро, Постелем и Россигнолем показывают, что продолжительность жизни этих рыб составляет более 5 лет.
Сардинелла круглая имеет наибольшее распространение в шельфовых водах тропической части Западной Африки от м. Кабо-Бланко до Анголы. На всем этом огромном пространстве существует несколько локальных стад сардинеллы. Е. Постель отмечает пять основных районов, где наиболее часто встречается этот вид. Три из них изобилуют сардинеллой: это район Рио-де-Оро - Сенегал, Берег Слоновой Кости - Того и Среднее Конго - Ангола. Два других, в которых, по мнению Постеля, сардинелла круглая встречается в меньших количествах, расположены у Гвинеи - Либерии и Нигерии -Габона. По морфометрическим признакам сардинелла всех этих районов разделяется на две расы, одна из которых обитает у северо-западного побережья Африки (до Гвинейского залива), а другая у Среднего Конго и Анголы. Большую часть жизни сардинелла круглая проводит вблизи берега, на глубинах 20—50 м. В этих же местах происходит ее нерест. Половой зрелости она достигает на втором году жизни. По данным Фюрнестина, сардинелла круглая обитает в водах с температурой на поверхности более 18°. Общий диапозон температур (от поверхности до дна), при которых можно встретить эту рыбу, изменяется от 15 до 30°. Промысел сардинеллы ведется как донными тралами, так и пелагическими орудиями лова. Она идет на изготовление деликатесных консервов, а также используется в качестве наживки на тунцеловном промысле.

Рис.3.1.2. Сардинела круглая (Sardinella aurita)
Из других видов сардинеллы у западного побережья Африки широко распространена плоская сардинелла Sardinella eba, которая отличается от круглой сардинеллы более высоким уплощенным телом и отсутствием желтой полоски на боках. Этот вид обитает преимущественно вблизи берега, в траловых уловах встречается редко. Рыбаки прибрежных стран ловят эту рыбу кошельковыми неводами. По вкусовым качествам она мало чем отличается от сардинеллы круглой.
Сардина марокканская является расой Sardina pilchardus Walbaum (рис. 3.1.3.). Жаберная крышка у нее радиально исчерчена, в спинном плавнике 13 лучей, в анальном — 15—16, в среднем продольном ряду чешуи 39—40; чешуя легко спадает. Верхнечелюстная кость достигает вертикали глаза, но не доходит до края зрачка. Зубов нет. Жаберные тычинки длинные. От европейской сардины отличается более коротким телом и длинной головой. Брюшные плавники и анальное отверстие у нее расположены ближе к хвосту. Количество позвонков у марокканской сардины колеблется от 49 до 52, чаще всего «их бывает 50. По Фюрнестину, марокканская сардина созревает в более ранние сроки, чем европейская. Начало развития половых желез у нее начинается в возрасте 8 месяцев, а первый нерест — в 12—16 месяцев

Рис.3.1.3. Сардина марокканская (Sardina pilchardus Walbaum)
И Восточной Атлантике марокканская сардина распространена от Канарских о-вов до Ла-Манша. Имеет очень большое промысловое значение. У западного побережья Африки к большом количестве вылавливается кошельковыми неводами в районе Марокко. Преобладающее большинство в уловах составляет сардина возрастом от одного до трех лет, длиной от 11 до 17 см. Промышляют ее преднерестовые и нерестовые скопления круглый год на расстоянии не далее 10— 15 миль от берега.
Нерест этой сардины происходит круглый год, достигая наибольшей интенсивности (по числу особей весной и в начале лета (март—июнь).
Существуют два района нереста марокканской сардины - северный и южный.
Северный - зимой располагается вблизи Касабланки и Федалы, на 5-15 миль от берега. Весной расширяется к северу до широты Лараге. Наибольшей интенсивности нерест в этом районе достигает в полосе воды от 2 до 8 миль от берега. Летом и осенью район нереста снова сужается, возвращаясь в свое зимнее положение (Касабланка, Федала).
Южный район зимой простирается от Могадора до Ифни. Наиболее массовый нерест сардины происходит между м. Гир и устьем р. Масса от 2 до 10 миль мористее береговой черты.
Весной, как и на севере, южный район имеет максимальную протяженность и простирается от города Сафи до м. Хуби. Летом он снова сужается, располагаясь между м. Гир и устьем р. Масса.
Южный район нереста более значителен, чем северный.
Большое влияние на сроки нереста марокканской сардины, на ее возрастное распределение по районам оказывает температура окружающей среды. Растянутость нереста, например, объясняется исключительно благоприятными температурными условиями, которые создаются у побережья Марокко почти во все сезоны года.
Упомянутые выше сроки наиболее массового нереста в годы, характеризующиеся повышенной температурой воды, наступают раньше и, наоборот, в холодные годы - позже. В возрастном распределении сардины подмечена следующая закономерность. Молодые, впервые нерестующиеся особи обитают в водах с температурой от 20 до 24°. Взрослая рыба придерживается более холодных вод с температурой 14—16°. В связи с этим в самом северном районе (к северу от м. Кантен) распространена преимущественно сардина в возрасте от 1 до 2 лет, так как здесь летом температура поверхности воды достигает максимальных величин (23—24°) и, независимо от сезона года, остается выше, чем в остальных районах побережья Марокко. Между м. Кантен и м. Гир обитает взрослая сардина возрастом от 2 до 4 лет, ибо здесь в течение года наблюдается наиболее низкая температура — от 14 до 16°.
И, наконец, у м. Гир и южнее -- до м. Хуби можно встретить как молодую, так и взрослую сардину возрастом от 1 до 4 лет.
Таким образом, намечается общая схема миграции, обусловленная возрастными изменениями в поведении марокканской сардины.
Молодая сардина, проведя первые стадии своего развития к северу от м. Кантен в водах с высокой температурой, совершает миграцию к более холодным водам, заполняющим район между м. Кантен и м. Гир. Из района вблизи м. Гир молодая сардина, достигнув определенного возраста, уходит в холодные воды, окаймляющие этот район как с севера, так и с юга.
Значительное влияние на перемещение скоплений сардины оказывают течения. Часто косяки ее переносятся ими на значительные расстояния. Сардина избегает районов с интенсивными течениями. Так как основной поток вод в районе Марокко имеет южное направление, то за выступающими мысами образуются циклонические круговороты вод с относительно слабыми течениями. В этих местах сардина находит наиболее благоприятные условия и концентрируется продолжительное время.
Вертикальные миграции марокканской сардины те же, что и у пелагических рыб: днем она небольшими стайками удерживается в придонных слоях воды, а с наступлением темноты поднимается к поверхности образуя мощное скопление. Причем, если эта концентрация сардины состоит из различных возрастных групп, то молодая рыба располагается выше, придерживаясь более теплых слоев воды, а взрослая держится ниже, где вода холоднее. Однако отмечены случаи, когда сардина ночью держалась мощными скоплениями в придонных слоях воды, а днем, разбиваясь на мелкие стайки, поднималась к поверхности, а иногда сардина почти в течение всех суток скапливалась у грунта и только утром на короткое время поднималась в верхние слои воды.
Сардина является прекрасным сырьем для изготовления деликатесных консервов.
Южноафриканская сардина (сардинопс) Sardinops ocellata (Pappe) (рис. 3.1.4.). Тело удлиненное, сжатое, слегка округлое. На брюшке мягкие килевые чешуйки. Чешуя легко спадает. У основания хвостового плавника имеются две увеличенные крыловидные чешуйки. Зубов нет. На боках, выше средней линии, расположен ряд пятен. В спинном плавнике 7—20 лучей, чаще - 18—19, в анальном 16—19 лучей, чаще 17—18.

Рис.3.1.4. Южноафриканская сардина (сардинопс) (Sardinops ocellata)
Длина сардинопса может достигать 30 см, но для промысловых уловов характерны экземпляры длиной от 18 до 24 см. Нерестовый период сардинопса растянут. Обитает эта рыба в поверхностных слоях воды. Питается в основном фитопланктоном. Распространена по юго-западному побережью Африки от п-ова Тигриш до м. Игольного. Используется для производства консервов, жира и кормовой муки.
Илиша Ilisha africana (Bloch) (рис.3.1.5.). Синоним Pellona africana Bleeker. Характерные признаки: сильно сжатое с боков тело, рот полуверхний, анальный плавник очень длинный, в нем 43 луча. Спинной плавник, в котором 12 или 13 лучей расположен позади маленького брюшного плавника. В среднем продольном ряду 40 чешуи от головы до основания хвостового плавника и 3 чешуи на последнем.

Рис 1.4.4.Илиша (Ilisha Africana)
Чешуя легко спадает. От горла до анального плавника имеются острые кетлевые чешуйки. Зубы мельчайшие. Длина до 25 см. Распространена илиша у западного побережья Африки в прибрежной зоне. Чаще всего встречается в распресненных водах вблизи устьев рек. Мясо съедобное, но икра в период размножения ядовита.
Лов сардины на свет
Сардины занимают большое место в мировом улове благодаря своей высокой численности. Особенно велик запас сардинопса.
На рис. 3.2.1 показаны районы обитания сардины. За последние годы значительное снижение мировых уловов сардины произошло за счет сардинопса, который в 1938 г. составлял 87,5% всего улова Тихого океана. Как известно, с 1942 г. промысел сардинопса в водах СССР, Кореи и на севере Японских островов из-за сокращения 'численности этой рыбы полностью прекратился и остался только на юге Японии. Значительно сократился промысел в северной части Тихого океана, где успешно вели лов США и Канада.

Рис. 3.2.1. Район обитания сардины:
1 - сардинопс; 2 - сардина; 3 - сардинелла.
В силу этого основной промысел сардины из района Тихого океана в настоящее время переместился в новые районы, расположенные главным образом у юго-западного побережья Африки, где добывается в основном также сардинопс.
В последнее время у западных берегов Африки начал успешно осваиваться промысел сардинеллы с крупных судов вооруженных придонными тралами.
Промысел сардины (сардина и сардинопс) ведется главным образом при помощи кошельковых неводов и лампар.
Часть сардины добывается с использованием света. В прибрежной зоне, таким образом ловят сардину Япония и все страны, расположенные в бассейне Средиземного моря.
Кошельковыми неводами сардину ловят на свет с наступлением темноты.
Бригада имеет на вооружении сейнер или мотобот мощностью 15-80л. с., баркас, кошельковый невод и мелкие суда или шлюпки от одного до пяти единиц с источниками света.
Кошельковый невод длиной 130-500 м ( в зависимости от судна) и высотой в зависимости от района работы (30 м при лове в прибрежном районе и 60 м в открытом море) и изготавливается из дели с размером ячеёй 5 - 7 мм в сливной и предсливной части, а в осносвной и бежной частях (до18 мм).
Источники света подвешены на кронштейне в кормовой части. Обычное число ламп - три, но встречаются лодки с одной, двумя, четырьмя, и большим количеством ламп. Лампы располагаются от воды на 0,5 - 0,6 м. и выше.
Лодки с зажженными лампами становятся на якорях. Расстояние между лодками от 100 до 150 м, а иногда и более.
В зависимости от концентрации рыбы продолжительность свечения ламп колеблется от 1 до 2 час. За это время сардина собирается в освещенной зоне в непосредственной близости от лампы. Для того чтобы за один замет кошелькового невода получить большой улов, лодки, кроме одной, снимаются с якоря и медленно движутся к той лодке, у которой скопление сардины наибольшее. Сведя лодки в одно место, на них тушат одну за другой лампы, оставляя со светом в зоне облова только одну лодку. Спустя 10—15 мин, когда отдельные скопления соединятся в единый косяк, с моторного судна начинается обмет кошельковым неводом. Чтобы не распугать рыбу, при обмете кошельковым неводом ворота невода располагают со стороны кормы лодки с источником света (рис. 3.2.2.). Одна лодка остается при судне для контроля и поддержания пятного крыла невода. Лодка со светом из зоны облова кошелькового невода выводится после частичной подсушки невода. Процесс кошелькования невода и выливка рыбы в зависимости от улова занимают от 1,5 до 2,5 час. В настоящее время при лове сардины применяют световую трассу.
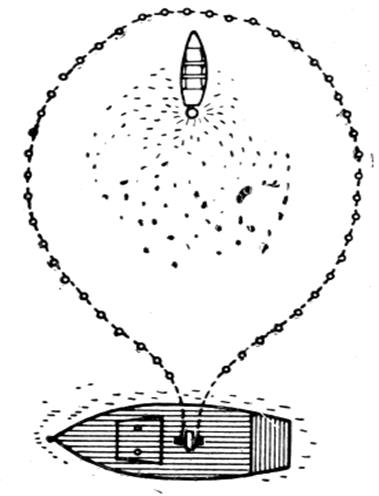
Рис 3.2.2.Расположение лодки с источником света в первый момент кошелькования
Техника лова в этом случае сводится к следующему. Судно на малом ходу вытравливает электрический кабель длиной 650-1200 м с прикрепленными к нему поводцами длиной 1,5 м. Поводцы отстоят на 100-150 м один от другого. На концах поводцов крепятся электролампочки мощностью 500 Вт каждая. После вытравливания кабеля в зависимости от промысловой обстановки судно ложится в дрейф или становится на якорь.
После того как образовалось достаточное промысловое скопление, к одной из крайней по трассе лампе подходит лодка с зажженным лампами. В момент подхода шлюпки с лампами, лампа на световой трассе выключается и скопление переводится к следующей лампе. Этот процесс повторяется до тех пор, пока вся скопившаяся на трассе рыба не будет сведена в единое скопление, удерживаемое в дальнейшем лодкой с лампами. После этого начинается замет кошелькового невода. Остальные операции производятся обычным способом. Новый комбинированный способ лова сардины имеет по сравнению со старым методом большие преимущества. Сардина быстрее накапливается в зоне подводного освещения, а улов за один замет доходит до 40 ц. Количество лодок с лампами сокращается до одного. Маневренность судна повышается, так как две оставшиеся лодки свободно могут быть размещены на судне. Облегчается труд и сокращается число рыбаков, улучшается техника безопасности, так как рыбаки во время работы находятся на судне. Применение подводного освещения в настоящее время на промысле сардины практикуется тунисскими рыбаками. Установлено, что лучшие результаты по привлечению рыбы дают лодки, оборудованные двумя лампами подводного освещения мощностью 500 вт.
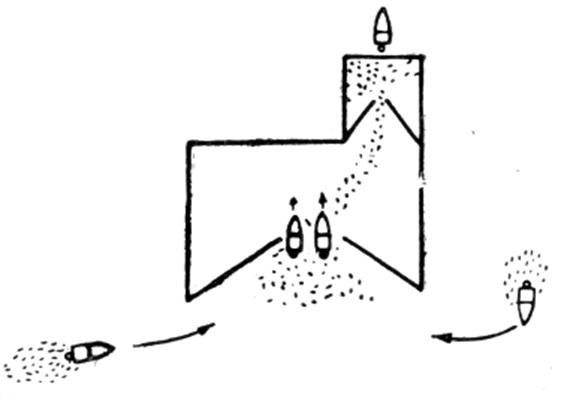
Рис. 3.2.3. Схема перевода рыбы во двор и (кассу) ловушки
Сардину на свет ловят не только кошельковым неводом, но и специальной ловушкой. На рис. 3.2.3. показана стационарная ловушка, называемая «сардинным заводом». По устройству и принципу действия эта ловушка сочетает в себе конструкцию обычного ставного невода и известного на Черном море кефалевого подъемного завода.
Сардинный завод служит только для удержания рыбы. Концентрируют рыбу у источника света около 10 лодок, оборудованных петромаксами. Сконцентрированная рыба при помощи медленно перемещаемого источника света заводится во двор завода, а затем ее переводят в «кассу». Для этого около кассы устанавливается на якоре лодка с включенным светом.
Особенно выгодно этот способ лова применять в тех районах, где по условиям работы невозможно использовать моторные суда.
В настоящее время сардину ловят на свет главным образом в бассейне Средиземного моря. И у берегов Японии.
Широко развит промысел ставриды и скумбрии на свет кошельковыми неводами у японцев. Он ведется в Восточно-Китайском море в районе о-вов Гото и Чечжудо. В этом виде промысла участвует сейнер, которому придаются три небольших судна, называемые «подсветчиками».
С этих судов опускаются «а глубину 50—55 м лампы подводного освещения и одновременно включается надводный свет. Собрав рыбу подводным светом, ее .постепенно поднимают и сводят в единый косяк. Обмет косяка производят сейнером, используя вместо шлюпки один из освободившихся подсветчиков. Второй подсветчик светит постоянно до тех пор, пока не выберут невод до сливной части. Третий подсветчик расправляет верхнюю подбору невода, препятствуя образованию углов.
Этим способом ловят главным образом мелкую ставриду, которая очень хорошо реагирует на свет. При лове крупной ставриды задача подсветчиков - обнаружить косяк и, если рыба реагирует на свет, задержать ее движение, пока сейнер не произведет замет. Если же рыба не реагирует на свет, подсветчик движется над косяком, показывая его направление, и сейнер делает замет, как на буй. Остальные операции те же, что и при лове мелкой ставриды. Размер кошелькового невода 300 х 800 м. После того как невод выбран до слипа, к сейнеру подходит транспортное судно и берет на борт верхнюю подбору невода. Улов выливают на транспортное судно при помощи специального сетного полотна, называемого «киталой». При таком способе добывается несколько миллионов центнеров ставриды и участвует большое количество судов.
Лов бортовым подхватом
Наибольшее распространение получили плоские бортовые подхваты, которые представляют собой прямоугольное сетное полотно, посаженное всеми кромками на подборы (рис. 3.3.1). Кромки основного сетного полотна могут иметь опушку из прочной нитки.
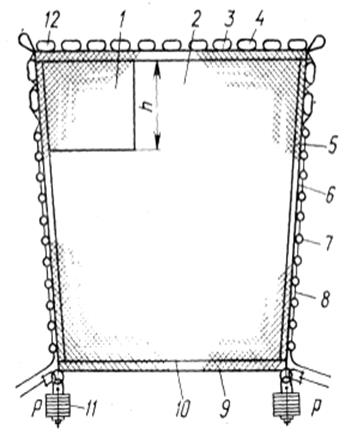
Рис.3.3.1. Плоский бортовой подхват:
1 – сливная часть; 2 – основное сетное полотно; 3 – верхняя подбора; 4 – наплав; 5 – боковая опушка; 6 – боковая подбора; 7 - стяжные кольца; 8 – стяжной трос; 9 – нижняя подбора; 10 – нижняя опушка; 11 – груз; 12 – верхняя опушка.
Иногда в одном из углов подхвата, примыкающем к верхней подборе, делают из более прочной дели слив, в котором концентрируется рыба перед выливкой улова.
Длина подхвата ограничена размерами свободного борта судна, например расстоянием между траловыми дугами, и редко превышает 20—30 м. Высоту подхвата принимают такой, чтобы при его подъеме захватить большую часть рыбы, сконцентрированной у рабочего борта судна. Обычно она колеблется от 15 до 30 м. Сетное полотно сажают на подборы так, чтобы образовался мешок. Однако подхваты со слишком большим выдуванием менее удобны в работе. В практике лова дальневосточной сайры глубина сетного мешка не превышает 15—25 м. Иногда подхват в посаженном виде имеет трапецеидальную форму в результате посадки сетного полотна по верхней подборе с большим посадочным коэффициентом, чем на нижняя.
Подборы подхвата изготовляют из капронового или комбинированного каната. В последнем случае сетное полотно иногда сажают на посадочный трос, который затем крепят бензелями к комбинированному канату. Верхнюю подбору, а иногда и прилегающие к ней участки боковых подбор оснащают наплавами из пенопласта. На участках боковых подбор, свободных от наплавов, крепят стяжные кольца, через которые пропускают стяжной трос. К нижней подборе подхвата прикрепляют грузила. Кроме того, к нижним углам подхвата крепят груза. Они обеспечивают во время дрейфа судна положение подхвата, близкое к вертикальному. Чем выше сопротивление подхвата при дрейфе, тем больше берут груза. На практике суммарная масса грузов достигает 800 - 1000 кг. Это достаточно для работы подхвата при силе ветра 5—6 баллов. К грузам крепят троса для их подъема на палубу судна.
Подхват устанавливают за бортом судна (рис. 3.3.2.). Его верхнюю подбору растягивают между двумя выстрелами длиной 10 - 15 м, сваренными из стальных труб. На ноке выстрела имеется четыре обуха для крепления оттяжек и блоков. Другой конец выстрела выполнен в виде вилки для крепления к штырю вертлюга на планшире или в других местах в зависимости от типа судна и его назначения. Выведенные за борт выстрелы раскрепляют горизонтальными и вертикальными оттяжками. Через блок на ноке выстрела пропущена оттяжка подхвата. Ваера от нижних углов выстрела проводят через направляющие блоки на барабаны лебедки.
Для подъема и спуска грузов используют блоки траловых дуг или специальные кран балки. Иногда плоские подхваты снабжают мотнёй для концентрации рыбы. Она облегчает выливку улова, хотя в целом затрудняет работу с подхватом.
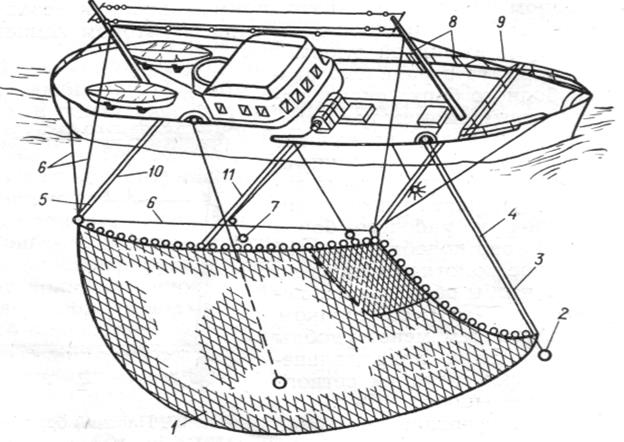
Рис. 3.3.2. Общий вид установки бортового подхвата для лова атлантической сардины:
1 - бортовой подхват; 2 - груз; 3 - стяжной трос; 4 - ваер; 5 - выстрел; 6 - стрела; 7 - концентрирующая лампа; 8 - кабели левого борта; 9 – крепление борта; 10 -оттяжка подхвата; 11 – бесконечный трос для установки концентрирующей лампы.
Рассмотренные конструкции подхватов и необходимое для работы с ними оборудование не совсем удобны в эксплуатации. При волнении возможен выход из строя выстрелов, оттяжек и других деталей под влиянием динамических нагрузок. Некоторые преимущества перед плоскими бортовыми имеют бортовые кольцевые подхваты (рис. 100).
Кольцевой подхват представляет собой плоскую сетную стенку трапецеидальной формы, снабженную по нижней подборе разъемными кольцами для стягивания нижней подборы.
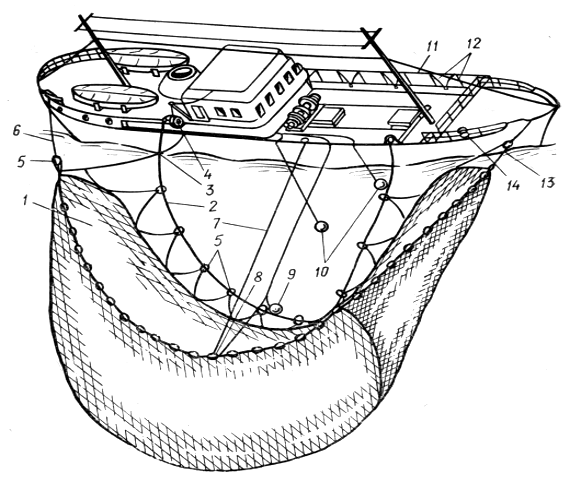
Рис. 3.3.3. Кольцевой бортовой подхват для лова атлантической сардины:
1 - кольцевой подхват; 2 - стяжной трос; 3 - бежной урез; 4 - силовой блок; 5 - разъемное кольцо; 6 - урез бежного крыла; 7 - фал для транспортировки концентрирующей лампы; 8 - кольцо для фала; 9 - концентрирующая лампа; 10 - белые лампы правого борта; 11- крепление ламп левого борта; 12 - кабели ламп левого борта; 13 - пятной урез; 14 - шпиль.
Кольцевой подхват имеет большие размеры, чем плоский, и позволяет облавливать большие по размеру световые скопления рыбы. Кроме того, он удобнее в работе при ветре более 3—5 баллов.
Размер ячеи сетного полотна и диаметр сетной нити у бортовых подхватов всех видов принимают из тех же соображений, что и в
сливе кошельковых неводов. Обычно сетное полотно подхвата освещено, но рассматривается на темном фоне, поэтому его окрашивают в темный цвет.
6.1.4. Лов кильки
Биология кильки
В Каспийском море обитает три вида килек (рис. 4.1.1): обыкновенная (Clupeonella delicatula caspia), анчоусовидная (С. engrauliformis), большеглазая (С. grimmi).
Обыкновенная килька была впервые обнаружена Богдановым в 1874 г., большеглазая - Кесслером в 1877 г. и анчоусовидная - Бородиным в 1904 г.
Кильки всех трех видов являются пелагическими стайными рыбами. Внешне они отличаются формой тела, окраской и другими признаками. Так, тело обыкновенной кильки в большей степени, чем у анчоусовидной, сжато с боков; спинка и верхняя часть головы пигментирована и имеют светло-зеленый оттенок. Брюшко заостренное с хорошо выраженным брюшным килем. Длина тела обыкновенной кильки меньше, чем большеглазой и анчоусовидной и составляет в Северном Каспии 8 см, в Южном 9 см.
Для анчоусовидной кильки характерно вальковатое удлиненное (прогонистое) тело; спина и верхняя часть головы пигментирована и имеют темно-синий цвет с зеленоватым оттенком. Анчоусовидная килька крупнее других килек. Длина ее тела составляет в среднем 10,5-11 см. У большеглазой кильки тело сильно сжато с боков; спина и верхняя часть головы темного цвета с фиолетовым оттенком. Глаза и голова по сравнению с обыкновенной и анчоусовидной килькой заметно крупнее; грудные плавники более длинные, а на брюшке имеется хорошо выраженный брюшной киль. Нижняя челюсть в большей степени, чем у других видов, выдвинута вперед. Большеглазая килька крупнее обыкновенной, но несколько мельче анчоусовидной.
После нереста килька отходит от прибрежной зоны в глубь моря, а с мелководий Северного Каспия, за небольшим исключением, откочевывает на юг. На протяжении лета и осени она держится во многих районах Среднего и Южного Каспия над глубинами 20 м и выше.
Аичоусовидная килька, в отличие от обыкновенной, обитает сравнительно далеко от берегов. Зимой основная масса взрослой кильки держится в Южном Каспии, в западной и восточной струях течения, а также на северных и южных склонах Апшеронского порога. В это время она придерживается глубоких слоев воды (40—80 м).
Обыкновенная килька является преимущественно обитателем прибрежной зоны моря. Зимой она держится в Среднем и Южном Каспии, главным образом в зоне с глубинами 30—100м.
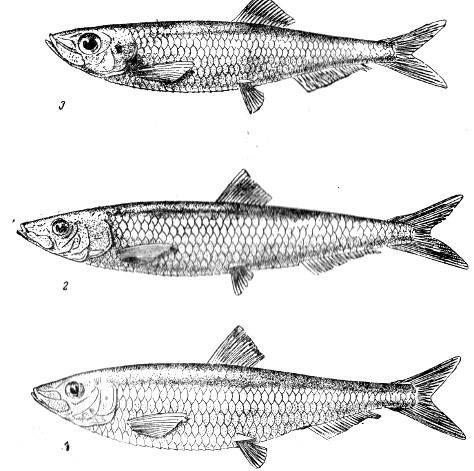
Рис. 4.1.1. Виды каспийских килек:
1 - обыкновенная; 2 – анчоусовидная; 3 – большеглазая.
Ранней весной килька поднимается в верхние слои и мигрирует на нерест. Большая часть кильки, мечущей икру в Северном Каспии, передвигается в этот район вдоль западного и восточного берегов моря. Нерест начинается во второй половине апреля и растягивается почти на весь май. Килька, нерестящаяся в Среднем и Южном Каспии с марта по май, подходит в прибрежную зону с глубинами до 2 м. После нереста килька отходит от прибрежной зоны в глубь моря, а с мелководий Северного Каспия, за небольшим исключением, откочевывает на юг. На протяжении лета и осени она держится во многих районах Среднего и Южного Каспия над глубинами 20 м и выше.
Аичоусовидная килька, в отличие от обыкновенной, обитает сравнительно далеко от берегов. Зимой основная масса взрослой кильки держится в Южном Каспии, в западной и восточной струях течения, а также на северных и южных склонах Апшеронского порога. В это время она придерживается глубоких слоев воды (40 - 80 м).
Весенние миграции анчоусовидвой кильки начинаются в апреле и мае (при поздней весне - даже в июне) и продолжаются до конца августа. В это время косяки перемещаются из Южного Каспия в Средний как по западному, так и по восточному побережью. Обычно к июню килька достигает северной части Среднего Каспия, где и образуются промысловые скопления ее. Часть косяков, зимовавших в самых южных участках Южного Каспия, во время весенней миграции не выходит за его пределы.
Летом и осенью (с июня по октябрь) значительная часть анчоусовидной кильки держится в Среднем Каспии главным образом в зоне глубин 50 м и выше. Нерест анчоусовидной кильки растянут и происходит летом и осенью. Обратная миграция ее из Среднего Каспия в Южный начинается в октябре и продолжается до конца декабря. Таким образом, продолжительность пребывания кильки и ее промысел в Среднем Каспии составляют около 5 месяцев.
Большеглазая килька, в отличие от двух других видов, является наиболее глубоководной формой, приспособившейся к обитанию на больших глубинах. В зимнее время она обитает главным образом в Южном Каспии над глубинами более 60— 90 м. Весной косяки этой кильки перемещаются в Средний Каспий, где образуют обычно наибольшие концентрации в районах с глубинами 80—150 м. Летом в Среднем Каспии большеглазая килька держится довольно разреженно, главным образом в зоне свалов, и лишь в отдельных случаях образует значительные скопления. Осенью, по мере охлаждения вод Среднего Каспия, большеглазая килька отходит на зимовку в воды Южного и южной части Среднего Каспия.
Кильки всех трех видов совершают суточные и сезонные вертикальные миграции, изучение которых представляет не только биологический интерес, по и имеет большое практическое значение для правильной организации килечного промысла. Знание горизонтов с максимальной концентрацией кильки, на которые следует опускать конусную сеть или всасывающий шланг рыбонасоса в различные сезоны, обусловливает успешное проведение лова.
Вертикальное распределение анчоусовидной кильки, которая составляет основную часть промысловых уловов. Весной (в мае) основная масса кильки сосредоточивается в верхнем прогретом слое воды, на глубине 10—15 м, но часть ее еще остается на глубине 30—40 м. Летом, по мере прогрева верхних слоев, килька опускается глубже и держится на глубине 10—30 м, где температура колеблется в пределах 9—19° и где располагается зона температурного скачка. Осенью (в октябре), с понижением температуры в верхних слоях и повышением в нижних, килька опускается глубже и распределяется по вертикали в более широких пределах (15—50 м), чем летом. Зимой килька опускается в еще более глубокие слои, чем осенью, и сосредатачивается в массе на глубине 30 – 80 м, а иногда и ниже.
Днем, как показывают записи эхолота, килька держится в различных горизонтах, в местах скоплений зоопланктона, и опускается до глубин 50 - 60 м. Такое распределение кильки, вероятно, связано с условиями ее питания (поиском пищи). Ночью она, как и зоопланктон, поднимается выше и распределяется в зоне с температурами 9 - 19°. Указанный диапазон температур, по видимому, является наиболее благоприятным для усвоения пищи и обмена веществ.
Подобное распределение кильки в слое температурного скачка может быть отменено с других позиций. В. А. Красильников указывает, что этот слой имеет повышенную плотность воды. По этой причине в нем собирается планктон, водоросли, мелкие морские организмы и т. п. Можно предположить, что по тем же причинам в слое температурного скачка собирается и килька. Моряки-подводники часто называют его слоем жидкого грунта; в нем легко уравновешивается подводная лодка.
Весной слой оптимальной для кильки температуры воды, а следовательно, и наибольшие скопления ее располагаются ближе к поверхности воды; летом же в связи с повышением температуры в поверхностных слоях килька опускаем глубже в зону оптимальных температур.
Осенью, с охлаждением поверхностных слоев воды высота слоя с оптимальными температурами увеличивается, увеличивается по вертикали и слой с наибольшей концентрацией кильки.
В зимний период температура воды понижается еще больше и сравнивается по вертикали. Слой температурного скачка в этот период исчезает, килька становится менее подвижной и отпускается в более глубокие слои.
Для оценки перспектив развития килечного промысла на Каспии необходимо знать состояние сырьевых запасов кильки и возможную величину ее вылова.
Проведенные «в 1952—1955 гг. КаспНИРО и промысловой разведкой промыслово-биологические съемки, распределения кильки показывают, что запасы ее весьма велики. Такое же мнение высказано всеми исследователями, занимавшимися изучением численности кильки. Даже по минимальной оценке запас был определен в 5 млн. ц.
Определение запасов кильки специальный вопрос, который выходит за пределы данной работы; можно лишь отметить, что лучшим подтверждением благополучного состояния запасов является непрерывный рост вылова с начала организации килечного промысла на Каспии, особенно за последнее пятилетие (табл. 1). До 1951 г. промыслом осваивалась главным образом обыкновенная килька. С 1951 г., в связи с освоением нового способа лова конусными сетями на электросвет, уловы кильки резко увеличились за счет использования анчоусовидной кильки.
Однако, несмотря на значительный рост добычи, можно с достаточной уверенностью утверждать, что запасы каспийской кильки до настоящего времени используются недостаточно. Увеличение добычи должно происходить за счет освоения запасов всех трех видов кильки, особенно анчоусовидной, так как запасы ее наиболее многочисленны.
В настоящее время районы, освоенные промыслом, занимают сравнительно небольшую акваторию и расположены вблизи приемных баз и портов-убежищ.
Лов кильки на электросвет производится в летне-осенний период с июня по октябрь, а иногда и позже, в районах Кзыл-Узень, Иралиево и Махачкалы; в зимние месяцы в районе Кианлы. Круглогодичный промысел производится в районе острова Куринский Камень. Несомненно, что по мере дальнейшего технического оснащения килечного промысла, промысловые районы будут расширены.
Техника лова кильки на свет
Дата: 2018-12-21, просмотров: 1523.