Согласно современным представлениям, в основе возникновения аритмий лежит нарушение образования или проведения импульса (потен-циала действия). Однако чаще всего аритмии возникают при участии обо-их механизмов (рис. 1).

Патогенез нарушений ритма сердца
| Нарушение образования импульса | Нарушение проведения импульса | ||||||||||||||
|
|
|
| |||||||||||||
| Изменение | Аномальный | Триггерная | Блокада | Повторный | Сверхнор- | ||||||||||
| нормального | автоматизм | активность | проведения | вход | мальное | ||||||||||
| автоматизма | импульса |
| импульса |
| проведение | ||||||||||
|
| (re-entry) |
| импульса | ||||||||||||

Рис. 1. Патогенез сердечных аритмий
Нарушения образования импульса. Нарушение образования им-пульса может быть обусловлено нарушением автоматизма и повышением возбудимости кардиомиоцитов.
Автоматизм (автоматия) — способность кардиомиоцитов спонтанно генерировать потенциал действия. Способностью к автоматизму обладают все атипичные кардиомиоциты (клетки проводящей системы сердца), но не обладают рабочие кардиомиоциты (клетки сократительного миокарда).
Возбудимость — свойство клеток возбудимых тканей воспринимать действие раздражителя и реагировать на него реакцией возбуждения. Возбудимость сердечной мышцы выражается в способности генерировать потенциал действия в ответ на раздражение.
В норме автоматизм проявляет только синоатриальный узел (СА-узел, узел Киса–Флека), являющийся номотопным (т. е. нормально расположен-ным) водителем ритма. Частота генерации импульсов клетками СА-узла
в покое у взрослого человека составляет 60–90 в мин. СА-узел называется истинным водителем ритма, или пейсмекером 1-го порядка (от англ. pacemaker — водитель ритма). Остальные структуры проводящей системы сердца (атриовентрикулярный узел, пучок Гиса, ножки пучка Гиса, волокна Пуркинье) также способны спонтанно генерировать импульсы, однако соб-ственная частота разрядов клеток этих отделов мала. Она тем ниже, чем дальше от пейсмекера 1-го порядка расположены клетки (градиент автома-
6
тии). Благодаря этому в нормальных условиях потенциал действия в этих клетках проводящей системы сердца возникает в результате прихода воз-буждения от более часто разряжающихся верхних отделов (клетки СА-узла), и их собственный автоматизм «не успевает» проявиться.
Таким образом, нижележащие структуры проводящей системы серд-ца проявляют автоматизм лишь при нарушении поступления импульсов от СА-узла и поэтому называются латентными (скрытыми, потенциаль-ными) водителями ритма.
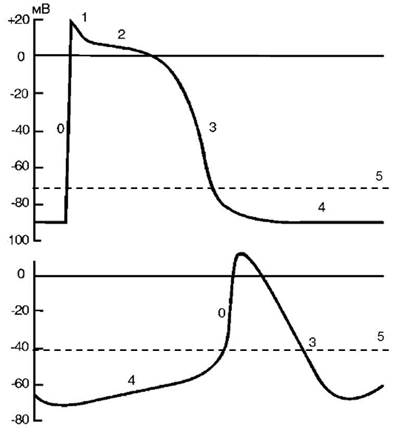
Изменение нормального автоматизма . Как известно, в основе про-цесса автоматизма лежит медленная спонтанная диастолическая деполя-ризация (рис. 2), постепенно уменьшающая мембранный потенциал до критического уровня (порог возбуждения), с которого начинается быстрая деполяризация мембраны (фаза 0 потенциала действия).
а
б
Рис. 2. Потенциал действия:
а — рабочий (типичный) кардиомиоцит; б — клетка синоатриального узла (атипич-ный кардиомиоцит): 0 — фаза быстрой деполяризации; 1 — фаза ранней быстрой ре-поляризации; 2 — фаза медленной реполяризации (фаза «плато»); 3 — фаза быстрой реполяризации; 4 — фаза потенциала покоя; 5 — порог возбуждения
Изменения нормального автоматизма сердца приводят к возникнове-нию синусовых аритмий. На продолжительность спонтанной диастоличе-
7
ской деполяризации и, следовательно, частоту генерации импульсов клет-ками СА-узла оказывают влияние следующие механизмы:
1. Скорость спонтанной диастолической деполяризации (наиболее важный). При ее возрастании порог возбуждения достигается быстрее
и происходит учащение синусового ритма (тахикардия). Замедление спонтанной диастолической деполяризации ведет к замедлению синусо-вого ритма (брадикардия).
2. Величина потенциала покоя клеток СА-узла. Если величина по-
тенциала покоя становится более отрицательной (например, в результате гиперполяризации мембраны при действии ацетилхолина), то требуется больше времени для достижения порога возбуждения (при условии, что скорость спонтанной диастолической деполяризации остается неизмен-ной) — возникает брадикардия. Если потенциал покоя клеток СА-узла становится менее отрицательным, то, соответственно, требуется меньше времени для достижения порога возбуждения — развивается тахикардия.
3. Изменение порога возбуждения. Более отрицательное значение величины порога возбуждения клеток СА-узла способствует учащению синусового ритма, а менее отрицательное значение — его урежению.
Возможны и различные комбинации трех основных электрофизиоло-гических механизмов, регулирующих автоматизм СА-узла.
Аномальный автоматизм . В норме ритм сердечных сокращений задают клетки СА-узла, все остальные клетки проводящей системы серд-ца разряжаются, как и рабочий миокард, под действием распространяю-щегося возбуждения. Потенциал действия в них возникает под влиянием токов от возбужденных участков миокарда до того, как в результате их собственной медленной спонтанной диастолической деполяризации их мембранный потенциал достигнет порога возбуждения.
Аномальный автоматизм — это появление пейсмекерной активности в клетках сердца, не являющихся в норме водителями сердечного ритма (т. е. они становятся водителями ритма сердца вместо клеток СА-узла).
Если по той или иной причине возбуждение СА-узла не возникает либо не может перейти на предсердие вследствие нарушения проводимо-сти, роль водителя ритма берет на себя атриовентрикулярный узел (АВ-узел, узел Ашоффа–Тавары) — пейсмекер 2-го порядка (частота ге-нерации импульсов 40–60 в мин). Если проведение возбуждения от пред-сердий к желудочкам полностью нарушено, то желудочки сокращаются в ритме пейсмекера 3-го порядка (частота генерации импульсов менее 40 в мин), расположенного в проводящей системе желудочков.
СА-узел называют номотопным (нормально расположенным) води-телем ритма, а очаги возбуждения в остальных отделах проводящей системы сердца — гетеротопными (ненормально расположенными). Эк-
8
топический водитель ритма — источник сердечных импульсов, располо-женный в любом месте миокарда помимо СА-узла.
Сокращение сердца, вызванное импульсом из гетеротопного очага возбуждения, называют замещающим, а последовательность сокраще-ний — замещающим ритмом (например, АВ-ритм). Такие ритмы выпол-няют защитную функцию, поддерживая относительно высокую частоту сердечных сокращений (ЧСС) при выраженном замедлении импульсации из СА-узла.
Гетеротопные очаги возбуждения могут определять ЧСС в случае, если их скорость генерации импульсов (соответственно, частота) будет больше, чем у СА-узла. Например, на фоне высокой концентрации ка-техоламинов может усиливаться автоматизм клеток нижележащих отде-лов проводящей системы сердца, и, если частота их деполяризации ока-зывается больше частоты генерации импульсов клетками СА-узла, это приводит к появлению эктопического ритма (например, экстрасистолия). Такие эктопические ритмы могут возникать при ишемии миокарда, гипо-ксии, при электролитных нарушениях и т. д.
При поражении миокарда (например, ишемии и др.) рабочие кардио-миоциты могут приобретать патологическую способность к автоматизму. Если скорость спонтанной диастолической деполяризации таких кардио-миоцитов превышает частоту генерации импульсов клетками СА-узла, то эти клетки устанавливают собственную ЧСС, становясь эктопическим водителем ритма.
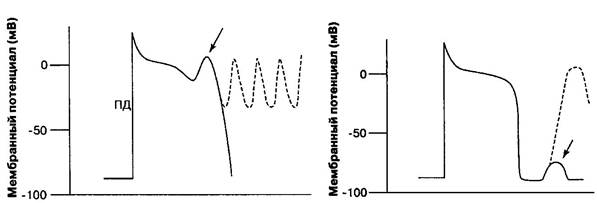
Триггерная активность . Повышение возбудимости кардиомиоцитов наиболее часто обусловливает возникновение аритмий по механизму триггерной активности (наведенной, пусковой, от англ. trigger — спуско-вой крючок). Электрофизиологической основой триггерной активности (триггерного автоматизма) являются ранние и поздние постдеполяриза-ции (рис. 3).
ПД
а б
Рис. 3. Потенциал действия: триггерная активность:
а — ранняя постдеполяризация; б — поздняя постдеполяризация
9
Ранняя постдеполяризация — это преждевременная деполяризация кардиомиоцитов, которая появляется в фазу реполяризации потенциала действия, когда потенциал мембраны еще не достиг величины потенциала покоя (рис. 3). Условиями возникновения ранних постдеполяризаций яв-ляются удлинение фазы реполяризации потенциала действия и брадикардия.
При замедлении реполяризации и, соответственно, увеличении об-щей продолжительности потенциала действия может возникнуть прежде-временная спонтанная деполяризация в тот момент, когда процесс репо-ляризации еще не завершился. При уменьшении частоты основного ритма сердца (брадикардии) происходит постепенное возрастание амплитуды ранних надпороговых колебаний мембранного потенциала, что при до-стижении величины порога возбуждения может вызывать образование нового потенциала действия еще до завершения исходного.
Этот преждевременный потенциал действия рассматривается как триггерный (наведенный), поскольку он обязан своим возникновением ранней постдеполяризации, исходящей от основного потенциала дей-ствия. В свою очередь, второй (наведенный) потенциал действия за счет своей ранней постдеполяризации может вызвать третий, тоже триггерный потенциал действия и т. д.
Если источник триггерной активности находится в желудочках, то может развиться желудочковая экстрасистолия или полиморфная (парок-сизмальная) желудочковая тахикардия (желудочковая тахикардия типа «пируэт» — torsade de pointes). Электрокардиографическая картина поли-морфной желудочковой тахикардии представлена на рис. 4 — желудочко-вые комплексы как бы вьются вокруг изоэлектрической оси (здесь и далее ЭКГ приведены во II стандартном отведении, скорость записи — 25 мм/с).
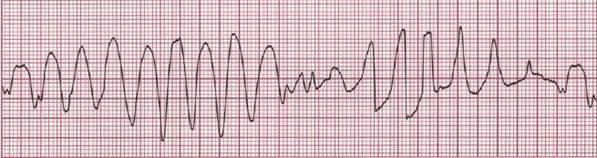
Рис. 4. Полиморфная желудочковая тахикардия
Возникновению ранних постдеполяризаций способствуют: гиперка-техоламинемия, гипокалиемия, ацидоз, ишемия.
Поздняя постдеполяризация — это преждевременная деполяризация кардиомиоцитов, которая появляется сразу после завершения фазы репо-ляризации, т. е. когда электрический заряд мембраны соответствует по-тенциалу покоя (рис. 3).
10
Подпороговые колебания мембранного потенциала, которые в норме могут присутствовать, но никогда себя не проявляют, при патологических состояниях, сопровождающихся увеличением внутриклеточной концен-трации Са2+ в кардиомиоцитах, могут возрастать по амплитуде, достигая порога возбуждения (рис. 3). Повышение внутриклеточной концентрации Са2+ вызывает активацию неселективных ионных каналов, обеспечиваю-щих усиленное поступление катионов (главным образом Na+) из внекле-точной среды в кардиомиоцит. В результате отрицательный заряд внут-ренней поверхности мембраны уменьшается, достигая величины порога возбуждения, и возникает серия преждевременных потенциалов действия.
Поздние постдеполяризации часто появляются при инфаркте миокарда, могут возникнуть под действием сердечных гликозидов, катехоламинов.
Нарушения проведения импульса. Проводимость — способность кардиомиоцитов проводить импульсы возбуждения. Существует три ос-новных типа нарушений проводимости: замедление или блокада проведе-ния, повторный вход импульса (механизм re-entry), сверхнормальное про-ведение.
Замедление или блокада проведения импульса . Распространение импульса возбуждения блокируется, если он достигает участка миокарда, неспособного к возбуждению. Чаще всего проведение импульса наруша-ется, если он достигает кардиомиоцитов, находящихся в рефрактерной фазе. В этом случае говорят о функциональной блокаде. Распространение следующего импульса возбуждения может не нарушаться. Блокада, вы-званная барьером в виде рубцовой ткани, замещающей кардиомиоциты, называется фиксированной блокадой проведения импульса.
Причиной замедления проведения импульса возбуждения или его блокады нередко бывает снижение количества потенциалзависимых Na+-каналов клеток, которым в норме присуще свойство быстрой деполя-ризации (например, рабочие кардиомиоциты, волокна Пуркинье). Суще-ствует прямая зависимость между количеством потенциалзависимых Na+-каналов и величиной потенциала покоя. Так, при уменьшении мем-бранного потенциала покоя рабочего кардиомиоцита до уровня –50 мВ (в норме –90 мВ) инактивируется около половины всех Na+-каналов.
В этом случае возбуждение и проведение импульса становятся невозмож-ными (например, в зоне ишемии миокарда).
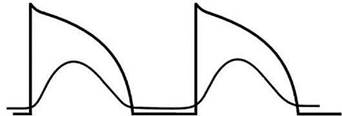
В определенных случаях даже при значительном уменьшении потен-циала покоя проведение импульса возбуждения, правда, существенно за-медленное, сохраняется. Такое проведение осуществляется медленными Са2+-каналами и медленными Na+-каналами, которые устойчивы к сниже-нию потенциала покоя (рис. 5).
В рабочих кардиомиоцитах существуют только быстрые Na+-каналы, но в условиях ишемии миокарда одна половина этих каналов инактивиру-
11
ется, а другая половина может превратиться в аномальные «медленные» Na+-каналы. Таким образом, «быстрые» клетки превращаются в «медлен-ные» кардиомиоциты, при прохождении через которые импульс возбуж-дения может замедлить свое распространение или блокироваться.
1
2
Рис. 5. Влияние острой ишемии миокарда на потенциал действия рабочего (типично-го) кардиомиоцита:
1 — нормальный потенциал действия; 2 — «медленный» потенциал действия
Повторный вход импульса (механизм re - entry ). Этим термином обозначают явление, при котором электрический импульс, совершая дви-жение по замкнутому кругу (петле, кольцу) в миокарде, возвращается к месту своего возникновения (circus movement).
Различают macro re - entry (макрориентри) и micro re - entry (микрори-ентри). При таком делении учитывают размеры круга (петли), в котором осуществляется повторный вход импульса.
В норме импульсы из СА-узла строго упорядоченно распространя-ются по сердцу, вызывая деполяризацию всех волокон миокарда, после чего затухают. Каждый участок миокарда и проводящей системы деполя-ризуются под действием одного импульса только один раз (в силу ре-фрактерности кардиомиоцитов).
Для развития механизма re-entry необходимы следующие условия:
– существование 2 каналов проведения импульса, разделенных меж-ду собой функционально или анатомически;
– односторонняя блокада проведения импульса;
– наличие потенциально замкнутой петли движения импульса (например, в местах разветвления волокон проводящей системы, зонах контактов окончаний волокон Пуркинье с кардиомиоцитами);
– замедление проведения импульса по цепи re-entry (замедление скорости проведения импульса, так что ни в одной точке петли волна воз-буждения не встречается с зоной рефрактерности).
Соблюдение этих условий возможно в том случае, если соседние клетки миокарда будут различаться по скорости проведения импульсов
и продолжительности рефрактерного периода (например, функционально гетерогенный участок при ишемии миокарда).
На рис. 6 показано распространение волны возбуждения в норме. Импульс достигает точки х и далее следует по двум параллельным путям (α и β) в нижележащие отделы проводящей системы. Скорость проведе-
12
ния и рефрактерность путей α и β одинаковы, поэтому две волны возбуж-дения одновременно приходят в дистальный участок проводящей систе-мы, где гасят друг друга (затухание импульса).
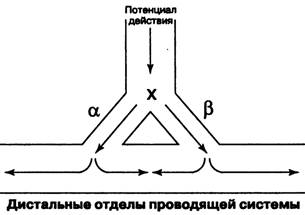
Рис. 6. Проведение импульса в норме (конечные волокна Пуркинье)
На рис. 7 показано, что происходит, если проведение импульса по одному из путей блокировано. В этом случае прохождение импульса воз-буждения из верхних в нижние отделы проводящей системы возможно только по пути α (рис. 7, а). Распространяясь, импульс может достигнуть конечной точки пути β (у). Если проводимость дистальных отделов пути β тоже нарушена, то импульс продолжает распространяться в нижележащие участки. Однако иногда импульс способен проводиться по пути β ретро-градно (вверх), и это явление — одно из обязательных условий реализа-ции механизма re-entry (рис. 7, б).
Нарушение, при котором импульс распространяется по проводящим путям только ретроградно, называют односторонней блокадой. Односто-ронняя блокада проведения импульса наблюдается в участках миокарда, где соседние клетки имеют разный рефрактерный период (например, при ишемии миокарда).
Импульс, распространяющийся ретроградно по пути β, может вновь достигнуть точки х. Если в этот момент путь α находится в фазе реполя-ризации, то повторная волна возбуждения затухает в точке х (рис. 7, б).
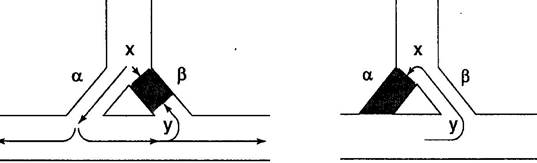
а б
Рис. 7. Механизм повторного входа импульса (re-entry). Односторонняя блокада
13
Если скорость ретроградного проведения импульса по пути β мень-ше, чем в норме, то в этом случае время, за которое повторная волна воз-буждения проводится по пути β, достаточно для полной реполяризации пути α (рис. 8). Достигнув точки х, импульс вновь активирует путь α, за-мыкая круг, по которому постоянно циркулирует волна возбуждения. Этот круг может существовать неопределенно долгое время, при этом каждое прохождение импульса активирует дистальные отделы проводя-щей системы, после чего волна возбуждения распространяется по всему миокарду, вызывая тахиаритмии.
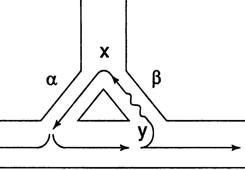
Рис. 8. Механизм повторного входа импульса (re-entry). Замедление ретроградного проведения импульса
Таким образом, сущность механизма re-entry заключается в том, что импульс повторно входит в участок миокарда или проводящей системы, создавая циркуляцию волны возбуждения.
Сверхнормальное проведение . Возникает в случае, если импульс возбуждения из СА-узла быстрее достигает желудочков по дополнитель-ным путям предсердно-желудочкового проведения. К таким путям отно-сятся пучок Кента (правый и левый), Джеймса и некоторые другие. Пра-вый пучок Кента соединяет правое предсердие с правой ножкой пучка Гиса, а левый пучок — левое предсердие с левой ножкой пучка Гиса. Пу-чок Джеймса соединяет предсердия с нижней частью АВ-узла. По этим дополнительным проводящим путям импульсы возбуждения из СА-узла достигают части желудочков раньше импульсов, которые идут обычным путем через АВ-узел. Это приводит к преждевременной активации части желудочков, другая их часть возбуждается позже импульсами, проходя-щими через АВ-узел. При этом происходит взаимное наложение прово-димых импульсов возбуждения и возникает тахиаритмия (например, син-дром Вольфа–Паркинсона–Уайта).
14
Дата: 2019-05-28, просмотров: 425.