Функции нервной системы
1. Регуляция деятельности всех органов и систем, эндокринная и иммунная регуляция, поддержание внутренней среды организма
2. Взаимодействие организма с внешней средой, анализ различных видов внешних раздражителей
3. Ответная реакция организма на различные виды воздействий, направленная на адаптацию к окружающей среде
4. Контроль за положением тела в пространстве, регуляция всех видов двигательных актов.
5. Осуществление всех видов психической деятельности (сознание, эмоциональное реагирование, речь, память, гнозис, праксис, мышление, интеллект, особенности человеческой личности).
Нервная клетка
Нервную ткань составляют нервные клетки (нейроны) с их отростками и клетки глии. Сосуды и оболочки мозга имеют мезодермальное происхождение.
Нейрон является основной структурной и функциональной единицей нервной системы и состоит из тела (сома) и отростков. Как правило имеются один длинный маловетвящийся центральный отросток –аксон и короткие периферические, имеющие много ответвлений дендриты.
Только в головном мозге насчитывается около 10 млрд нейронов.
Основные функции нейрона:
-проведение нервного импульса
-синтез белков, липидов, углеводов
-синтез нейромедиаторов в некоторых случаях гормонов.
Нейроны делятся на сенсорные (чувствительные), моторные (двигательные) и ассоциативные, а по характеру вырабатываемого в них медиатора дифференцируются на холинергические, серотонинергические, адреналинергические, ГАМКергические, допаминергические и.т.п.
Тело клетки и ее отростки покрывает непрерывная сдвоенная мембрана (невролемма), выполняющая разграничительную и транспортную функцию. Благодаря мембране осуществляется пассивный транспорт воды и низкомолекулярных веществ, а также работа ионных насосов для поддержания внутри нейрона постоянного мембранного потенциала покоя и возникновения возбуждающего потенциала действия с резким изменением проницаемости для содержащих биоэлектрический заряд ионов, что определяет возникновение нервного импульса.
В теле нейрона располагается цитоплазма и ядро с ядрышком, содержащие ДНК и РНК. В ядре сконцентрирован генетически аппарат клетки.
Комплекс Гольджи участвует в синтезе углеводных полимеров и гормонов. Митохондрии содержат дыхательные ферменты, обеспечивают процесс окислительного фосфорилирования и участия в гликолизе, их основная функция синтез богатой энергией АТФ. Лизосомы, содержащие гидролитические ферменты и расщепляют для последующей биоутилизации биологические макромолекулы внутриклеточного и внеклеточного происхождения. Эндоплазматическая сеть с гранулами РНК и рибосомами ответственны за синтез белков.
Нервное волокно-в большинстве случаев аксон нервной клетки, состоящий из осевого цилиндра, покрытого миелиновой оболочкой. Миелиновая оболочка состоит из спиралевидно окружающих осевой цилиндр аксона слоев, при этом толщина аксона, в основном, зависит от толщины миелиновой оболочки и составляет от 0,3 до 20 мкм. Миелиновая оболочка аксона продуцируется из оболочек прилегающих к нейрону глиальных клеток, в центральной нервной системе –олигодендроцитов, в периферической нервной системе-шванновских клеток (леммоцитов). Между фрагментами миелиновой оболочки образуются просветы, лишенные миелина-перехваты Ранвье. Скорость проведения по волокну нервного импульса прямо пропорциональна толщине его миелиновой оболочки и варьирует от 0,6 до 20 м/с. Наибольшей скоростью проведения обладают толстые миелинизированные волокна. Тонкие слабомиелинизованные нервные волокна проводят импульс с существенно меньшей скоростью.
Для нейрона характерна исключительно высокая активность метаболических процессов. Биосинтез большинства макромолекул осуществляется в теле нервной клетки. Отсюда они распространяются по аксону посредством ортоградного аксонального тока (аксональный транспорт). Благодаря аксотоку по аксону транспортируются молекулы белка, гликопротеиды, фосфолипиды, некоторые ферменты, необходимые для поддержания метаболизма. Продукты, происходящих в аксоне процессов метаболизма посредством ретроградного аксонального тока перемещаются в тело клетки, где подвергаются дальнейшему разрушению ферментами лизосом до составных эелементов, используемых при формировании вновь синтезируемых в теле нейрона макромолекул.
Нервная система способна преобразовывать воздействие внешних и внутренних раздражающих факторов в энергию нервного импульса, имеющую электрическую природу.
При этом нервный импульс перемещается всегда в одном направлении: от дендритов к телу клетки, а затем по аксону.
Нейроглия
Глиальных клеток в нервной системе примерно в 10 раз больше, они не имеют аксонов и сохраняют способность к делению в постнатальном периоде. Выделяют астроглию, олигодендроглию и эпендимную глию.
Астроглия состоит из астроцитов, которые участвуя в формировании гематоэнцефалического барьера (ГЭБ), выполняют важную роль в обеспечении метаболических процессов в нервных клетках. Астроциты располагаясь между нейронами и капиллярами, избирательно пропускают в нейрон биологически активные вещества и продукты обмена. Было доказано участие астроцитов в метаболизме глутамата и ГАМК, которые являются соответственно возбуждающими и тормозными медиаторами. При нарушении иммунных процессов в нервной ткани астроциты приобретают свойства иммуноэффекторных клеток, экспрессирующих ряд антигенов и иммуномодуляторов, которые формируют в мозговой ткани иммунный ответ, что играет роль в патогенезе аутоиммунных демиелинизирующих заболеваний нервной системы.
Олигодендроглия сконцентрирована в белом веществе мозга, формирует миелиновую оболочку нервных волокон. На периферии функции олигодендроцитов выполняют шванновские клетки (леммоциты).
Эпендимная глия выстилает расположенные внутри мозга ликворные пути (желудочки и водопровод мозга, а также центральный канал спинного мозга) и выполняет, таким образом, барьерную функцию.
Глия выполняет, главным образом, опорную функцию, формируя своеобразный каркас, в ячейках которого находятся тела нейронов, обеспечивает относительную изоляцию и обособление нейронов, поддержание ионного баланса, выполняет аккумулирующую функцию, поглощая выделяемые нейронами при их «зарядке» и «разрядке» свободные электролиты. Глиальные элементы, взаимодействуя с нейронами, обеспечивает транспортные функции, обладают способностью к накоплению РНК, белка, дыхательных ферментов, обеспечивают возможность поглощения и накопления запасов медиаторов с последующим их высвобождением. Глия (в основном леммоциты) участвует в процессах дегенерации и регенерации нервной ткани. Есть мнение о том, что глия является местом хранения закодированной информации, что обеспечивает, так называемую долговременную память.
Разрастание глиальной ткани при повреждении мозга ведет к формированию мозгового рубца. Установлено также, что астроциты участвуют в поддержании ионного баланса и, следовательно, влияют на генерацию нервного импульса, регулируя уровни Na+ и K+ вокруг клетки. Считается, что формирование глиальных рубцов нарушает забуферивание K+ вокруг нервной клетки, что приводит к снижению порога возбудимости нейрона и эпилептическим разрядам.
Таким образом, изменение состояния и функции глии может влиять на развитие многих физиологических процессов и заболеваний нервной системы.
Гематоэнцефалический барьер
Эндотелий капилляров и расположенных в мозговых желудочках сосудистых сплетений вместе с элементами глиальной ткани (главным образом астроцитами) составляют гематоэнцефалический барьер (ГЭБ).
Контакты между эндотелиальными клетками достаточно плотные, промежутки между ними, как правило, меньше 1,5 нм, что служит препятствием для большинства молекул, в частности белковых. К тому же между капилляром и нейроном располагается астроцитарная глия. Эти структуры служат своеобразным фильтром, обеспечивающим относительную защиту нервных клеток и их отростков.
Таким образом, ГЭБ регулирует поступление в цереброспинальную жидкость и нервную ткань циркулирующих в крови веществ, обеспечивая относительное постоянство внутренней среды мозга. ГЭБ защищает мозг и цереброспинальную жидкость от всевозможных случайных эндогенных и экзогенных факторов, которые, попав в кровяное русло, могли бы оказать на нервную систему неблагоприятное воздействие.
При патологических состояниях проницаемость ГЭБ может увеличиваться, что обусловливает проникновение в нервную систему веществ, приводящих к нарушению в ней гомеостаза и развитию различных патологических состояний (отек, набухание, аутоиммунные процессы и др.). Проницаемость ГЭБ играет роль и в лечении патологических процессов нервной системы, т.к. для достижения терапевтического эффекта необходимо учитывать возможность проникновения того или иного лекарственного средства через ГЭБ.
Строение нервной системы
Нервная система анатомически состоит из центрального и периферического отделов. Кроме того выделяют соматическую (анимальную) и вегетативную (автономную) нервную системы. Соматическая нервная система регулирует произвольные движения поперечно-полосатой мускулатуры скелета, языка, гортани, глотки, обеспечивает общую чувствительность и связь с внешней средой. Вегетативная нервная система в большей степени влияет на функции гладких мышц внутренних органов, сосудов, сердечную деятельность, секреторную функцию железистой ткани, в том числе и эндокринных желез, регулирует обменные процессы во всех органах и тканях. Вегетативная нервная система не подчиняется воле человека, а потому считается автономной и подразделяется на симпатический и парасимпатический отделы, действующие функционально антагонистически.
Центральная нервная система это расположенный в полости черепа головной мозг и находящийся в позвоночном канале спинной мозг.
К периферической нервной системе относятся парные спинномозговые и черепные нервы с их корешками, их ветви, образующие нервные сплетения и периферические нервы, нервные окончания и ганглии. Периферические нервы в зависимости от того, из каких волокон (чувствительных либо двигательных) они состоят, подразделяются на чувствительные, двигательные и смешанные. Тела нейронов, отростки которых составляют чувствительные нервы, лежат в нервных узлах вне мозга. Тела двигательных нейронов лежат в передних рогах спинного мозга или двигательных ядрах головного мозга.
Макроскопически головной и спинной мозг состоят из серого и белого вещества. Серое вещество представляет из себя скопление тел нервных клеток. Отдельные ограниченные скопления серого вещества в головном мозге носят названия ядер, в спинном мозге соответственно называются рогами.
Поверхность головного мозга –серая, благодаря наличию множества тел нейронов. К серому веществу головного мозга относят кору больших полушарий, базальные (подкорковые) ганглии, ядра зрительных бугров и гипоталамуса, скопления тел нейронов в стволе головного мозга (ядра черепных нервов, ретикулярная формация, бугорки четверохолмия, красные ядра, черная субстанция). Серое вещество спинного мозга представлено скоплениями тел нейронов центральной части сегмента спинного мозга и локализовано в передних, боковых и задних рогах спинного мозга.
Белое вещество-это отростки нервных клеток (аксоны) с миелиновыми оболочками, составляющие проводящие пути головного и спинного мозга.
ПРОВОДНИКИ ЧУВСТВИТЕЛЬНОСТИ
Пути поверхностной и глубокой чувствительности анатомически различны.
Весь путь поверхностной чувствительности состоит из четырех нейронов и носит название tractus ganglio- spino- thalamo- corticalis.
Первый нейрон –это клетки периферических, чувствительных нейронов заложенные в межпозвоночных спинальных узлах (ганглиях); их периферические отростки — чувствительные волокна периферических нервов — проводят импульс с периферии от нервных чувствительных окончаний (рецепторов).
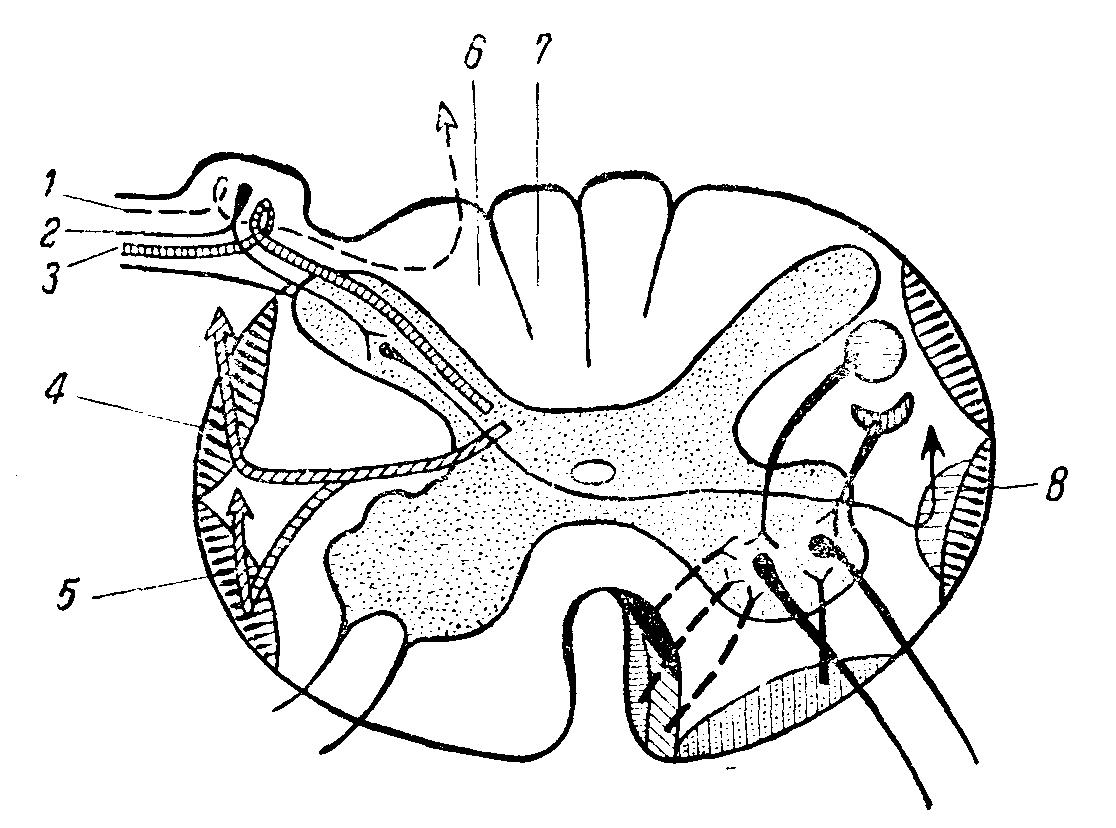
Рис. 1. Поперечный срез спинного мозга. [Формирование центростремительных проводников из волокон задних чувствительных корешков и подход волокон центробежных путей к клеткам передних рогов.
1 — волокна суставно-мышечной, вибрационной и тактильной чувствительности; 2 — волокна болевой и температурной чувствительности; 3 — волокна мозжечковых проприоцепторов; 4 — дорсальный спино-церебеллярный путь; 5 — вентральный спино-церебеллярный путь; 6 — пучок Бурдаха; 7 — пучок Голля; 8 — спино-таламический путь.
Центральный отросток клеток межпозвоночных узлов проводит импульс в составе волокон заднего чувствительного корешка в спинной мозг, а именно в клетки заднего рога сегмента спинного мозга, где располагается второй нейрон.
Из клеток заднего рога возникает основной спинномозговой проводящий путь поверхностной чувствительности именуемый спиноталамическим трактом, волокна которого через переднюю серую спайку постепенно переходят на противоположную сторону и устремляются в центральном направлении вверх в составе боковых столбов спинного мозга, достигая таламуса.
Следует учесть, что в спиноталамическом тракте волокна, идущие из нижних сегментов спинного мозга располагаются более латерально (снаружи) относительно волокон, идущих из верхних сегментов, что важно для топической диагностики поражений спинного мозга. В соответствии с данным законом экцентрического расположения длинных проводников при наличии патологических процессов внутри спинного мозга (например интрамедуллярная опухоль) первоначально будут сдавливаться внутренние (медиальные) волокна спиноталамического тракта иннервирующие вышележащие сегменты и соответственно уровень чувствительных расстройств будет спускаться сверху вниз (нисходящий тип). Напротив, при экстрамедуллярных процессах (когда патологический очаг располагается вне спинного мозга) в первую очередь сдавлению подвергаются проводники наружних отделов спинного мозга от нижележащих сегментов. В этом случае уровень чувствительных нарушений будет постепенно подниматься от нижних конечностей по восходящему типу.
Достигая продолговатого мозга спиноталамический тракт располагается в боковых его отделах. На уровне варолиева моста подходит к медиальной петле, объединяющей в себя пути поверхностной и глубокой чувствительности и далее заканчивается в ядрах зрительного бугра (таламус), где расположены тела третьих нейронов.
От нейронов таламуса начинается tractus thalamo- corticalis, волокна которого через заднюю треть заднего бедра внутренней капсулы, а далее в составе corona radiatа идут в заднюю центральную извилину, где располагается четвертый нейрон.
Распределение чувствительных волокон в коре представлено в порядке обратном расположению частей тела. Верхние отделы задней центральной извилины заняты центрами чувствительности для нижних конечностей, средние отделы -для верхних конечностей, в нижней части находятся центры чувствительности лица. Кроме того чувствительные волокна оканчиваются в коре верхней теменной дольки, которая наряду с задней центральной извилиной является корковым отделом кожного анализатора.
Таким образом, вследствие перекреста волокон второго нейрона в сегментах спинного мозга, импульсы поверхностной (болевой, температурной и частично тактильной) чувствительности передаются в заднюю центральную извилину с противоположной стороны тела. Соответственно, поражение первого или второго нейрона до перекреста (уровень периферического нерва, межпозвоночного ганглия, корешка, заднего рога) вызывает расстройства чувствительности на стороне поражения. Если же пострадали волокна второго нейрона после перекреста или волокна третьего и четвертого нейронов (уровень проводников спинного мозга, мозговой ствол, зрительные бугры, внутренняя капсула, кора), то расстройства чувствительности наблюдаются на стороне противоположной очагу.
Пути глубокой чувствительности также состоят из четырех нейронов tractus ganglio- bulbo- thalamo- corticalis
Клетки первых нейронов заложены в межпозвоночных ганглиях. Их периферические отростки заканчиваются в рецепторах глубоких тканей (мышцы, связки, сухожилия, надкостница). Центральные же отростки через задние корешки не заходя в серое вещество спинного мозга, вступают прямо в задний столб своей стороны спинного мозга и в составе пучков Голля и Бурдаха поднимаются, не прерываясь, вверх до продолговатого мозга, где и заканчиваются в ядрах задних столбов или ядрах Голля и Бурдаха, где располагаются клетки вторых нейронов. Следовательно, первые, или периферические, нейроны обеспечивают проведение импульса от периферии до продолговатого мозга, причем проводники следуют все время по своей стороне спинного мозга.
Следует помнить, что в медиально расположенном пучке Голля оказываются проводники от нижних конечностей, в латеральном же пучке Бурдаха — проводники от более высоких сегментов, главным образом верхних конечностей.
От ядер Голля и Бурдаха начинаются волокна вторых нейронов, которые в продолговатом мозге совершают перекрест, на уровне варолиева моста вливаются в медиальную петлю, где присоединяются к волокнам болевой и температурной чувствительности и заканчиваются в зрительных буграх (таламус).
Слияние обоих чувствительных путей — tractus spino-thalamicus и bulbo-thalamicus, начавшись в продолговатом мозге, завершается окончательно только в мосту. Общий чувствительный путь мозгового ствола, составленный двумя названными пучками, носит наименование медиальной петли — lemniscus medialis. Медиальная петля представляет собой собрание волокон вторых нейронов всех видов чувствительности противоположной стороны тела (перекрест волокон болевой и температурной чувствительности происходит последовательно по сегментам на всем протяжении спинного мозга в передней серой спайке, а волокон суставно-мышечного и тактильного чувства — в продолговатом мозге). Кроме того в мозговом стволе к медиальной петле присоединяются чувствительные проводники от черепных нервов (языкоглоточный, блуждающий, тройничный).
Нервные волокна медиальной петли завершаются в ядрах таламуса, где расположены тела третьих нейронов глубокой чувствительности. Проводники несущие импульсы из ядер зрительных бугров, минуя внутреннюю капсулу и corona radiata подходят к корковым центрам глубокой чувствительности, представленных в задней центральной извилине и теменной доле (четвертый нейрон).
Благодаря корковым центрам осуществляется высший анализ и синтез проприоцептивных раздражений, позволяющих ощущать движение, перемену положения частей тела.
Локализация функций в коре
Функции коры разнообразны и многогранны. В целом, это высшая регуляция всех нервных процессов, начиная с сознания, осуществления произвольных двигательных актов, анализа всех видов чувствительных стимулов, включая специфическую чувствительность (зрение, слух, обоняние, вкус) и заканчивая сложноорганизованной и сложноподчиненной мозговой деятельностью, сочетающей в себе множество более простых нервных аналитико-синтетических актов именуемой высшей нервной деятельностью.
Четко локализовать функции в коре достаточно сложно в связи с большим количеством ассоциативных зон перекрытия, многочисленных связей коры как с нижележащими подчиненными отделами нервной системы, так и с другими корковыми зонами своего и противоположного полушария мозга.
Самые простые функции коры больших полушарий представлены в так называемых проекционных зонах, которые относительно отчетливо связаны с определенной областью периферической (двигательной или чувствительной) иннервации. Совокупность периферической (рецепторной или эффекторной) и центральной (корковой) проекционных зон с проводящими путями, соединяющими их воедино называется анализатором.
Двигательные зоны для произвольных движений мускулатуры противоположной стороны тела расположены в передней центральной извилине.
Проекция для отдельных мышечных групп представлена здесь в порядке, обратном расположению их в теле: верхним отделом передней центральной извилины соответствует нога, средним отделам — рука и нижним отделам передней центральной извилины — лицо, язык, гортань и глотка.
Проекции поворота глаз и головы в противоположную сторону соответствует задний отдел средней лобной извилины.
Чувствительные зоны находятся в задней центральной извилине. Проекция кожных рецепторов аналогична соматотопическому представительству в передней центральной извилине: в верхнем отделе извилины представлена чувствительность нижней конечности, в среднем — руки и в нижнем — головы.
Зрительная зона расположена в затылочных долях, на внутренней поверхности полушарий, по краям и в глубине шпорной борозды. В каждом полушарии представлены противоположные поля зрения обоих глаз, причем область, расположенная над шпорной бороздой (клин), соответствует нижним, а область под ней (язычная извилина) — верхним квадрантам полей зрения.
Слуховая проекционная область коры находится в височных долях, в верхней височной извилине и в извилинах Гешля (на внутренней поверхности височной доли).
Обонятельная проекционная область расположена также в височных долях, главным образом в крючке парагипопокампальной извилины (uncus). Есть основания предполагать, что близко к обонятельным территориям расположены и вкусовые.
Все проекционные области коры являются двухсторонними, симметрично расположенными в каждом полушарии. Часть из них связана только с противоположной стороной (передняя и задняя центральные извилины, зона поворота глаз и головы, зрительная область). Корковые слуховые, обонятельные и вкусовые территории каждого полушария связаны с соответствующими рецепторными полями на периферии с обеих сторон (противоположной и своей).
Высшая нервная деятельность
Высшая нервная деятельность присуща в наибольшей степени человеку и является самой сложной и совершенной формой функционирования нервной системы. Высшие нервные функции физиологически и анатомически в наибольшей степени связаны с корой больших полушарий, к ним относят речь, праксис, гнозис, память, мышление, интеллект.
На основании физиологических и анатомических исследований и клинического опыта известно, что территории коры, связанные с осуществлением более сложных соподчиненных функций коры не являются строго локализованными, их развитие происходит в основном в постнатальном периоде; в этом имеется различие с проекционными территориями.
Высшая нервная деятельность осуществляется за счет бесчисленных связей, анатомическим субстратом которых являются ассоциационные и комиссуральные волокна белого вещества полушарий между соседними извилинами, а также связывающие между собою целые доли головного мозга и полушария. Эти пути проходят под корой головного мозга, в белом веществе полушарий, часто на значительной глубине, и составляют как бы магистрали, необходимые для обеспечения сложной, совместной, сочетанной деятельности обширных территорий, занимаемых нервными клетками коры.
Функции праксиса и гнозиса
Праксис-способность к осуществлению сложных, целенаправленных двигательных актов, которые создаются путем подражания, обучения и постоянной практики в течение индивидуальной жизни.
Гнозис способность узнавания предметов по тем или иным свойствам, сложная нервная функция, заключающаяся в постоянном анализе разнообразных чувствительных стимулов с возможностью интегрирования их в единый образ. Гнозис также как и праксис, совершенствуется в течении жизни на основании опыта и индивидуальной практики.
Праксис и гнозис как высшие формы проявления двигательной и чувствительной функции, свойственные особенно человеку, являются продуктом длительного развития как биологического, так и общественного.
Праксис неразрывно связан с гностическими функциями. Даже простое движение не может быть совершено без постоянной рецепции (понимания, узнавания) самого движения. Непрерывная информация со стороны рецепторов опорно-двигательного аппарата тем более необходима, когда осуществляется сложное и тонко дифференцированное движение. Высший анализ и синтез кинестетических раздражении осуществляется двигательным анализатором, работа которого и обеспечивает чрезвычайное разнообразие, точность и координированность сложных двигательных актов, выработанных в практике индивидуального опыта.
Наиболее ответственны за функции праксиса теменная и частично лобные доли левого полушария (у правшей). Нарушения праксиса называется апраксией. Апраксия является результатом поражения двигательного анализатора без явлений паралича или инкоординации движений. В результате нарушения синтеза и анализа теряются навыки сложных, целенаправленных, производственных действий. Правильное пользование предметами обихода, профессиональные рабочие процессы, смысловая жестикуляция, игра на музыкальном инструменте и т.д. у больного нарушаются.
Для возникновения апраксии необходимо поражение обширных зон левого полушария, а иногда и двухстороннее поражение. Наиболее отчетливые формы апраксии возникают все же при поражении левой теменной доли у правшей (нижней теменной дольки, надкраевой извилины). Своеобразные апраксические расстройства возможны и при очагах в лобных долях.
В равной мере и гнозию нельзя понимать в отрыве от праксии. В самом деле, в процессе познания внешнего мира имело громадное значение развитие человеческой руки как органа труда и речи, как средства общения.
Так же как для праксиса специальных четко ограниченных «гностических центров» не существует. Речь идет о сложных расстройствах функций, связанных с работой какого-либо анализатора. Нарушения гнозиса терминологически обозначается как агнозия. В зависимости от того какой вид гнозиса страдает различают астереогноз-нарушение узнавания предмета на ощупь, нарушения схемы тела (аутотопогнозию), слуховую, зрительную, обонятельную, вкусовую агнозию.
Простые функции поверхностного и суставно-мышечного гнозиса связаны с теменной долей входящей в систему кожного и кинестетического анализаторов. С этими зонами связана способность определять отдельные свойства предметов на ощупь (температура, форма, вес и.т.д.)
Дальнейший тонкий анализ и синтез кожных и кинестетических раздражении осуществляется в обширных областях обоих анализаторов, корковые отделы которых взаимно перекрываются, особенно в передних отделах теменной доли.
Более сложный целостный процесс узнавания предметов на ощупь именуется стереогнозом. Расстройство стереогнозии (астереогоноз) возникает при очагах в теменной доле, кзади от задней центральной извилины, при этом нарушаются тесные функциональные и анатомические связи с областью проекции движений руки, особенно кисти и пальцев. В таких случаях больной с закрытыми глазами не может узнавать предметы при ощупывании их противоположной очагу рукой.
Аутотопогнозия т.е. нарушения схемы тела возникает преимущественно при поражении теменной доли правого полушария у правшей. При этом человек теряет способность узнавать собственные части тела, различать правую и левую стороны. При поражении этой же корковой зоны может возникнуть анозогнозия, когда больной не осознает своего дефекта, например грубых двигательных нарушений.
Слуховая агнозия, т.е. нарушение способности различения предметов по характерным для них звукам (гул машин, шум воды, щебетание птиц, звон разбитого стекла и.т.п.) может наблюдаться при поражении височных долей, куда подходят афферентные слуховые проводники. Слуховая агнозия наблюдается редко; по-видимому, для возникновения ее необходимо обширное и притом двухстороннее поражение височных долей. Одним из частных видов слуховой агнозии является сенсорная афазия сущность которой заключается в неузнавании, непонимании словесной речи (агнозия звуковой речи, словесная агнозия).
Зрительная агнозия связана с поражением затылочных долей, их наружных поверхностей. Под зрительной гнозией подразумевается узнавание, различение предметов по их виду, внешним характеристикам. Частным проявлением зрительной агнозии является неспособность чтения (алексия).
Зрительная агнозия встречается в клинике редко; чаще она имеет временный, преходящий характер или является частичной; по-видимому, для возникновения ее необходимо выключение функций обеих затылочных долей (наружных поверхностей полушарий).
Обонятельная и вкусовая агнозия являются расстройствами сложного анализа и синтеза соответствующих раздражении проявляются в виде нарушении дифференциации запахов и вкусов. Такого вида расстройства возникают при патологических очагах на внутренней и нижней (базальной) поверхности височной доли.
Речь и ее расстройства
Речь является одной из поздних (филогенетически новых) функций больших полушарий. Речь является только человеческой функцией; цитоархитектонические поля, имеющие наибольшее отношение к речевой функции, существуют только у человека.
Человеческое мышление всегда является словесным. Словесное мышление и речь относятся ко второй сигнальной системе, свойственной лишь человеку. Возникновение речи было обусловлено эволюционным развитием и постепенной социализацией человека.
Различают два основных вида речи:
Экспрессивная или моторная речь, функция которой воспроизведение (продукция) словесных звуковых рядов. Часть двигательного анализатора специализировалась для осуществления этой функции. Понятно, что она сформировалась вблизи от проекционной области для движений губ, языка, гортани — в заднем отделе нижней лобной извилины (область Брока-область моторной речи), в доминантном полушарии (у правшей в левом).
Импрессивная или сенсорная речь- процесс различения, узнавания условных звуковых рядов — сигналов — по их звучанию и порядку расположения, различение отдельных звуков (фонем), сопоставление элементов ее с теми или иными представлениями, предметами, понятиями, т.е. узнавание, понимание устной речи.
Центр сенсорной речи соответственно близок по локализации к корковым проекционным зонам слухового анализатора и располагается в заднем отделе верхней височной извилины доминантного полушария (левого у правшей) (область Вернике).
Следовательно, моторный компонент речи (экспрессивной) является специальным видом праксиса; сенсорный — специальным видом слуховой гнозии. Обе речевые функции тесно взаимно связаны.
Функцию письма также можно рассматривать как часть экспрессивной речи и в то же время специальным видом праксиса т.е. воспроизведение словесных знаков и рядов в виде условно принятых графических обозначений. Соответственно зоны письма близки по расположению к центру Брока и корковой двигательной проекционной зоне для мышц кисти, это задние отделы средней лобной извилины.
Чтение же напротив является элементом сложного зрительного гнозиса и параллельно частью импрессивной речи. Поэтому в большей степени эта функция страдает при поражении угловой извилины теменной доли на границе с затылочной и височными долями (задний отдел верхней височной извилины), рядом с областью Вернике
Функция речи чрезвычайно сложна. Будучи проявлением высшей нервной деятельности и составной частью человеческого мышления, она не может быть узко локализованной в коре головного мозга. Клинический опыт показывает, во-первых, что разнообразие характера речевых расстройств очень велико; во-вторых, определение расположения очага поражения на основании только одних речевых расстройств достаточно затруднительно и нередко ошибочно.
В осуществлении сложнейших речевых функций участвуют не отдельные изолированные корковые области, а сложные функциональные системы, охватывающие обширные территории коры, но все же поражения разных участков коры вызывают качественно отличные нарушения речи.
Корковые нарушения речи называются афазиями. В отличие от анартрии (дизартрии), зависящей от паралича мускулатуры, участвующей в артикуляции, больной при афазии свободно двигает языком и губами, но утрачивает навыки (праксию) речевых движений.
Поражение области Вернике (в заднем отделе верхней височной извилины) приводит к развитию сенсорной (акустико-гностической) афазии. Понимание речи развивается у ребенка ранее других речевых функций, поэтому поражение области Вернике влечет за собой наиболее тяжкий ущерб, расстраивая функцию других, сопряженных с ней в деятельности отделов коры. Клинически сенсорная афазия проявляется как полное расстройство понимания речи, многоречивость (словесная окрошка), обилие замен звуков, искажающих речь больного на столько, что ее нельзя понять. При сенсорной афазии наблюдаются разной степени тяжести расстройства письма и чтения, выражающиеся в заменах звуков и букв. Человек с сенсорной афазией воспринимает свой родной язык как незнакомый, иностранный.
Страдания больного усугубляется тем, что он не понимает своей речи и ему кажется, что все окружающие сговорились не понимать его.
Поражение области Брока (задний отдел нижней лобной извилины) вызывает речевые расстройства по типу моторной эфферентной афазии являющейся по сути словесной апраксией. Центральным дефектом при этой форме афазии является патологическая инертность речевого процесса. Возникающие у больных паузы и повторы слов в предложении затрудняют или делают невозможными устную речь.
При попытках к называнию или к спонтанной речи нарушается возможность плавной смены слогов и слов, возникает застревание на слогах слова, повторение уже сказанных слов (персеверации). Нарушения повествовательной, вопросительной и побудительной речи является основным признаком. Больному трудно сказать просьбу, приказ, задание, пожелание, рассказать о чем-то важном и т.д. Больной что-то пытается сказать, иногда ругается, расстраивается потому что не может объяснить, что ему надо.
Другим весьма своеобразным видом афазии является амнестическая афазия — симптом поражения заднего отдела височной и нижнего отдела теменной доли. При этой форме афазии возникают трудности удерживания в слухоречевой памяти серии слов, воспринятой на слух речевой информации, что приводит к недостаткам понимания длинных, сложных речевых отрезков. Устная речь этих больных относительно сохранна, но им трудно называть предметы и действия, в их речи наблюдаются обильные словесные замены, многоречивость, больной как-бы забывает слова.
Семантическая афазия возникает при поражении теменно-затылочных областей доминантного по речи левого полушария. Для этой формы афазии характерно расстройство понимания сложных логико-грамматических оборотов (нарушение понимания смысла пословиц, предлогов и т.п). Этот дефект речи, как правило, сопровождается нарушениями счета. Больные плохо выполняют инструкции, включающие предлоги «над» - «под», наречия «слева от» - «справа от», «сверху» - «снизу». Понимание простых фраз, без сложных логико-грамматических оборотов, у них остается сохранным
Исследование афазий (очень краткое и ориентировочное) производится следующим образом.
1. Проверяется понимание обращенной к исследуемому речи путем предложения устных заданий, выполнения инструкций: поднять правую руку, левую заложить за спину; закрыть глаза и т.д. (сенсорная функция речи).
2. Изучается собственная речь исследуемого: свободно ли говорит, правильно ли, обладает ли достаточным запасом слов, нет ли парафазии, персевераций и т.д. (моторная функция речи). Предлагается повторение речи исследующего.
3. Проверяется способность понимания написанного путем предложения письменных заданий: сделать то или иное движение, взять тот или иной предмет и т.д. (функции чтения).
4. Проверяется, как исследуемый пишет: свободно или с затруднением, правильно ли построена фраза.
5. Предлагается с целью исключения амнестической афазии называть предметы.
Изолированно функция чтения, или лексии, нарушается при локализации очага в теменной доле, в угловой извилине. Расстройство понимания письменной речи (алексия) возникает при очагах на стыке височной доли (словесная гнозия) и затылочной доли (зрительная гнозия).
Функция письма или графии изолированно нарушается при поражении заднего отдела средней (второй) лобной извилины, рядом с проекционной областью поворота глаз и головы и движений руки в левом полушарии у правшей. Такое расположение понятно, поскольку процесс письма сопряжен с движением глаз по строчкам и осуществляется правой рукой. Графия является одним из видов сложной праксии; письменная речь заключается в начертании условных, соответствующих звукам значков (букв), составляющих в определенных сочетаниях слова и фразы. Утрата способности письма называется аграфией.
Симптомы раздражения долей больших полушарий- это патологическая локальная гипервозбудимость тех или иных корковых зон с искаженным усилением функции чаще пароксизмального характера, как правило, являющиеся «аурой» (предвестником) генерализованного эпилептического припадка.
Симптомы выпадения обусловлены органическим нарушением структуры или гибелью нервных клеток в области поражения с временным либо стойким выпадением или снижением неврологической двигательной либо чувствительной функции (негативные симптомы)
Поражение роландовой области, т. е. передней и задней центральных извилин, характеризуется выпадением двигательных или чувствительных функций на противоположной стороне тела. Очаги в области передней центральной извилины вызывают центральные параличи или парезы. Так как расположение двигательных клеток Беца здесь достаточно разбросано могут наблюдаться гемипарезы с явным преобладанием поражения руки, ноги или лица; не столь редки и чистые моноплегии. Преимущественное поражение ноги указывает на локализацию процесса в верхних отделах передней центральной извилины, руки — в среднем и лица и языка — в нижнем.
При раздражении области передней центральной извилины возникают судороги тонического и клонического характера в мускулатуре, иннервируемой раздраженным участком — припадки корковой, или джексоновской эпилепсии. Типичным для джексоновской эпилепсии является начало судорог с ограниченной группы мышц при ясном вначале сознании. Припадок может этим и ограничиться, но иногда «генерализуется», переходя постепенно в общий судорожный припадок с потерей сознания, иногда недержанием мочи, прикусом языка и т.д.
Очаги в области задней центральной извилины вызывают расстройства чувствительности на противоположной стороне тела. Для корковых поражений характерна утрата суставно-мышечного чувства, тактильного, болевого, температурного, локализации, тонкого различения характера и степени раздражения часто в сочетании с крайне неприятными, плохо локализуемыми, иррадиирующими ощущениями — гиперпатией.
Раздражение задней центральной извилины вызывает приступы «чувствительной» джексоновской эпилепсии — парестезии в соответствующих раздраженному отделу областях противоположной стороны тела. При распространении припадка парестезии «расплываются» по соседству. Чаще, однако, иррадиация раздражения происходит в направлении передней центральной извилины с развитием после чувствительной ауры тонико-клонических судорог.
Поражение лобных долей
В целом, функции лобных долей направлены на организацию, планирование и осуществление сложных двигательных актов, праксис, в том числе моторную речь, формирование поведения, характера, мышления и интеллекта, координацию движений и равновесие.
Поражение лобной доли в правом полушарии (у правшей) может не давать отчетливых явлений выпадения или раздражения функций.
Поражение задних отделов средней лобной извилины или области «произвольного» сочетанного поворота глаз и головы вызывает корковый паралич взора в противоположную очагу сторону. При этом возникает отклонение глазных яблок (часто и головы) в сторону очага. При раздражении же этой области возникают судорожные подергивания глаз и головы в противоположную пораженному полушарию сторону. Такого рода припадки также относятся к корковой, или джексоновской, эпилепсии.
К числу лобных симптомов, также наблюдающихся одинаково при поражении как левого, так и правого полушария, относится лобная атаксия, резче всего проявляющаяся в отношении туловища и вызывающая, главным образом, расстройства стояния и ходьбы. При тяжелых поражениях больной, не имея параличей, не в состоянии сохранять вертикальное положение тела и, будучи поставлен на ноги, падает (астазия) или, с трудом удерживаясь на ногах, не может ходить (абазия). Данные расстройства координации движений и равновесия обусловлены нарушением связей лобной коры с противоположным полушарием мозжечка.
Изредка при поражении лобных долей наблюдается своеобразное расстройство — хватательные феномены. «Автоматическим хватанием» называется непроизвольное схватывание рукой предмета при прикосновении им к ладони больного; «навязчивым хватанием» является непроизвольное стремление захватывать окружающие предметы или следовать рукой за приближаемым и отдаляемым от руки больного предметом с тенденцией к его захвату.
Расстройства психики при поражении лобных долей развиваются довольно часто и характеризуются изменениями поведения и характера в сочетании со снижением интеллекта (лобная деменция). Отмечаются апатия, снижение мотивации, торпидность психики, нарушается способность к организации и планированию, ослабление памяти и внимания, отсутствие критического отношения к своему состоянию, недооценка тяжести заболевания. Чаще, чем депрессивные, наблюдаются состояния эйфории, эгоцентричность, иногда агрессивность. Довольно типична неряшливость, неопрятность больного (мочеиспускание в палате на пол, в постель).
Нарушения контроля за тазовыми функциями достаточно характерны для лобных нарушений и чаще являются следствием нарушения организации, планирования акта мочеиспускания или дефекации, т.е. результат апраксии.
Нередко при поражении лобных долей отмечаются явления синдрома паркинсонизма (гипомимия, шаркающая походка, олигобрадикинезия) обусловленные разобщением двусторонней связей лобной коры с подкорковыми экстрапирамидными ядрами.
Моторная эфферентная афазия возникает только при поражении лобных долей доминантного полушария, (левого полушария у правшей), а именно при поражении области Брока- заднего отдела нижней лобной извилины. При этом расстройстве больной теряет способность говорить, речевая продукция снижается, запас слов ограничен, но сохраняется в основном способность понимания речи. Больной с моторной афазией говорит медленно, с трудом, допуская ошибки (аграмматизм), которые сразу же замечает. В более легких случаях больной владеет всем запасом слов, но говорит с задержкой, «спотыкаясь» на слогах.
Одновременно с утратой речи возникает и аграфия. Как моторная афазия, так и аграфия представляют собой особый вид апраксии.
Поражение теменной доли вызывает главным образом чувствительные расстройства. Рассмотрим сначала симптомы, возникающие в равной мере при поражении того или другого полушария.
Астереогнозия является результатом поражения как задней центральной извилины, так и области, расположенной кзади от нее. При этом больной теряет навыки узнавания, определения предмета на ощупь. Более редкой формой агнозии является аутотопоагнозия, или неузнавание частей собственного тела, искаженное восприятие его. При аутотопоагнозии возникают своеобразные нарушения представлений о собственном теле: больной путает правую сторону с левой, ощущает наличие третьей руки или ноги; путает последовательность пальцев и т.д. Аутотопоагнозия может возникать, по-видимому, при поражении как левой, так и правой теменных долей.
Аутотопоагнозия нередко сочетается с анозогнозией, при которой наблюдается отсутствие сознания своего дефекта, чаще всего — паралича. Больной утверждает, что он двигает парализованными конечностями, описывает, как он их поднял, согнул и т.д. Анозогнозия является результатом поражения правой теменной доли и наблюдается только при левосторонних гемиплегиях (у правшей).
Одним из расстройств, возникающих при поражении коры левой теменной доли, является апраксия. Наиболее отчетливые формы апраксии причем в обеих руках возникают при поражениях в области надкраевой извилины левого полушария: больной теряет способность производить сложные целенаправленные действия при отсутствии параличей и полной сохранности элементарных движений. В случаях резко выраженной апраксии человек становится совершенно беспомощным, теряя навыки привычных действий и способы пользования предметами. Так, больной не может самостоятельно одеться, застегнуться, принять пищу; путает последовательность действий, не доводит их до конца.
При поражении угловой извилины доминантного полушария (в левой теменной доле у правшей) наблюдается алексия — утрата способности расшифровки письменных знаков — понимания написанного. Одновременно расстраивается и способность письма. Больной обычно не обнаруживает полной аграфии, как при поражении второй лобной извилины, но делает ряд ошибок в письме, неправильно пишет слова, часто и буквы — вплоть до полной бессмысленности написанного. Алексия является одним из видов зрительной агнозии.
Раздражение верхней теменной дольки вызывает внезапный приступ парестезий сразу во всей противоположной половине тела, а не постепенное распространение парестезий, как при раздражении задней центральной извилины. Вслед за чувствительной аурой следует приступ судорог, возникающих сразу во всей мускулатуре противоположной стороны тела (или сразу общий судорожный припадок).
Поражение височной доли правого полушария (у правшей) может не давать отчетливых симптомов. Все же в большинстве случаев удается установить некоторые симптомы выпадения или раздражения, характерные для обоих полушарий.
Квадрантная гемианопсия (выпадение квадрантов полей зрения), является иногда одним из ранних симптомов поражения височной доли. Причина квадрантной гемианопсии лежит в неполном поражении волокон зрительной лучистости, проходящей через височную долю
Атаксия, резче выраженная в туловище, вызывает главным образом расстройства стояния и ходьбы и обусловлена разобщением связей височной коры с противоположным полушарием мозжечка. При выполнении координаторных проб может наблюдаться промахивание.
Довольно характерными для поражения височной доли следует считать приступы вестибулярно-коркового головокружения, сопровождающиеся ощущением нарушения пространственных соотношений больного с окружающими предметами (существует представительство вестибулярного аппарата в коре височной доли); нередки сочетания такого головокружения со слуховыми галлюцинациями (гулы, шумы, жужжание).
В отличие от поражений правого полушария, очаги в левой височной доле (у правшей) часто влекут за собой тяжкие расстройства речи.
Наиболее частым симптомом является сенсорная (акустико-гностическая) афазия, возникающая в результате поражения области Вернике, расположенной в заднем отделе верхней височной извилины. Больной утрачивает способность понимать речь. Слышимые слова и фразы не связываются с соответствующими им представлениями, понятиями или предметами; речь больному становится непонятной совершенно так же, как если бы с ним говорили на незнакомом ему языке. Установить с такого рода больным контакт с помощью речи чрезвычайно трудно: он не понимает, чего от него хотят, о чем его просят и что ему предлагают. Одновременно с этим расстраивается и собственная речь больного. В отличие от больного с моторной афазией, больные с поражением области Вернике могут говорить и часто отличаются излишней говорливостью и даже болтливостью, но речь становится неправильной; вместо нужного слова произносится ошибочно другое, заменяются буквы или неправильно расстанавливаются слога. В тяжелых случаях речь больного становится совершенно непонятной, представляя собой бессмысленный набор слов и слогов («салат из слов»). Больной с сенсорной афазией не понимает не только чужую речь, но и свою: отсюда ряд ошибок, неправильностей и т.д. (парафазия). Дефектов своей речи больной не замечает.
Поскольку чтение и письмо являются навыками, приобретенными позднее звуковой речи и поскольку функции лексии и графии находятся в тесной зависимости от функции понимания устной речи, постольку поражение области Вернике вызывает сопряженно не только нарушения правильности речи, но и расстройства чтения и письма (алексия и аграфия).
Характерными для поражении височной доли на границе с теменной в доминантном (левом у правшей) полушарии являются амнестическая и семантическая афазии описанные выше.
Симптомами раздражения височной коры являются слуховые, обонятельные и вкусовые галлюцинации, являющиеся иногда начальным симптомом («аурой») эпилептического припадка. Одностороннее разрушение этих чувствительных зон заметных расстройств слуха, обоняния и вкуса не вызывает (каждое полушарие связано со своими воспринимающими аппаратами на периферии с обеих сторон — своей и противоположной).
Еще более типичными для раздражения височной доли считаются возникающие после упомянутого вида чувствительных аур (или без них) своеобразные сноподобные состояния, когда все окружающее кажется ненастоящим, нереальным, но в то же время странно знакомым, как будто уже пережитым в прошлом («ощущение уже виденного»).
Поражение затылочной доли как области, связанной с функцией зрения, вызывает зрительные расстройства. Очаги в области шпорной борозды, расположенной на внутренней поверхности затылочной доли, вызывают выпадение противоположных полей зрения обоих глаз, что влечет за собой следующие расстройства.
Гомонимная (одноименная) гемианопсия-выпадение только левых или только правых полей зрения. При поражении области выше шпорной борозды — клина, возникает не полная, а лишь квадрантная гемианопсия противоположных нижних квадрантов; при поражении язычной извилины (ниже шпорной борозды) выпадают поля противоположных верхних квадрантов. Еще менее значительные очаги могут вызывать дефекты в виде островков в противоположных полях зрения, называемые скотомами. Скотомы обнаруживаются в обоих полях зрения и в одноименных квадрантах. Одним из ранних симптомов поражения зрительного анализатора является выпадение цветовых ощущений в противоположных полях зрения. Даже при обширных двухсторонних поражениях областей шпорной борозды редко наступает полная слепота: обычно сохраняется так называемое центральное, или макулярное, поле зрения обоих глаз.
При процессах, возникающих на широких территориях затылочных полей, в частности с поражением наружной поверхности левой затылочной доли (у правшей), может наблюдаться зрительная агнозия: больной не является слепым, он все видит, обходит препятствия, но теряет способность узнавать предметы по их виду, может быть агнозия только на цвета, может быть неузнавание лиц (больной не различает знакомых от незнакомых), домов на хорошо известной улице и т.д. Понятно, что вместе со зрительной агнозией развивается и неузнавание написанного (письма), т. е. нарушения чтения-алексия.
Одним из проявлений зрительной агнозии является и так называемая метаморфопсия, когда нарушается правильное узнавание очертаний контуров предметов: последние представляются исковерканными, изломанными, неправильными.
В возникновении такого рода расстройств имеет значение нарушение связей затылочной области с височной, имеющей отношение к восприятию и оценке пространственных отношений (корковый отдел вестибулярного анализатора).
К явлениям раздражения, которые могут наблюдаться при процессах в затылочной доле, относятся следующие.
Зрительные галлюцинации: при раздражении проекционного зрительного поля в области шпорной борозды возникают «простые» галлюцинации (фотомы), т. е. световые, иногда цветовые явления в виде искр, пламени, теней и т.д. Раздражение наружной поверхности затылочной доли дает «сложные» галлюцинации в виде фигур, предметов, иногда движущихся, часто извращенных и устрашающих (метаморфопсии), что говорит о близости очага к височной доле. Обычно зрительные галлюцинации являются началом развивающегося вслед за ними эпилептического припадка.
Поражения основания головного мозга характеризуются главным образом поражением черепных нервов.
Процессы в области передней черепной ямки вызывают при одностороннем поражении атрофию зрительного нерва с амаврозом или амблиопией на стороне поражения и утрату обоняния (аносмию) вследствие поражения обонятельных луковиц и трактов.
Симптомами поражения средней черепной ямки является выпадение функций глазодвигательного, отводящего и блокового нервов (III, VI и IV), вызывающее офтальмопарез или офтальмоплегию на стороне поражения, а также симптомы раздражения (невралгию) или нарушения проводимости тройничного нерва (расстройства чувствительности на лице).
Процессы на основании мозга в области ножки мозга вызывают альтернирующий паралич типа Вебера: на стороне поражения имеется паралич глазодвигательного нерва (птоз, мидриаз, расходящееся косоглазие) и на противоположной стороне — центральный паралич конечностей вследствие поражения в ножке мозга пирамидного пучка, перекрещивающегося ниже.
Поражения гипофиза, в частности опухоли его, сопровождаются битемпоральной гемианопсией (выпадение наружных полей зрения), нередко амблиопией или амаврозом из-за сдавления средних отделов хиазмы зрительных нервов и рядом эндокринных расстройств. В одних случаях может наблюдаться акромегалия (гигантский рост языка, носа, нижней челюсти, рук и ног), в других- патологическое ожирение, отсталость развития половых органов и вторичных половых признаков, ослабление половой функции. Наконец, в результате поражения инфундибулярной области могут возникнуть патологические жажда и голод (полидипсия и булимия), расстройства сна и т.д.
Поражения в области задней черепной ямки характеризуются поражением слухового и лицевого нервов (VIII и VII), иногда также отводящего и тройничного (VI и V) в случае локализации процесса в так называемом мосто-мозжечковом углу. При более каудальной локализации могут быть поражены блуждающий, языкоглоточный и подъязычный нервы (X, IX, XII). Мозжечковые расстройства при процессах в области задней черепной ямки отмечаются на стороне поражения, пирамидные — на противоположной, на своей или с обеих сторон.
Пути к мозжечку
Спиноцеребеллярные тракты основные афферентные проводящие пути, передающий нервные импульсы от рецепторов опорно-двигательного аппарата в мозжечок.
В мышцах, суставах, связках, сухожилиях и надкостнице расположены нервные окончания, проводники от которых идут в частности к мозжечку. Мозжечковая проприорецепция обеспечивает постоянную информацию о состоянии органов движения. Импульсы по афферентным волокнам периферического нерва достигают клеток, расположенных в межпозвоночном спинальном ганглии (I нейрон), откуда по соответствующим волокнам заднего чувствительного корешка проводятся в задний рог спинного мозга. Здесь первый (периферический) нейрон проприоцепторов мозжечка заканчивается, и импульс передается на клетки вторых нейронов. Центральные отростки вторых нейронов формируют задний и передний спиноцеребеллярные тракты (пучки Флексига и Говерса), проходящие в боковых столбах спинного мозга. Задний спиноцеребеллярный тракт или пучок Флексига не перекрещиваясь входит в мозжечок через нижние ножки мозжечка и заканчивается в его черве. Пучок Говерса дважды перекрещиваясь вступает в червь мозжечка в составе верхних ножек мозжечка. При поражении пучков Флексига и Говерса мозжечковые расстройства возникают на стороне поражения.
Кроме основных спиноцеребеллярных трактов мозжечок имеет многочисленные афферентные связи с другими структурами ЦНС, обеспечивающих равновесие и двигательную координацию.
Вестибуло-церебеллярный тракт обеспечивает связь вестибулярных ядер с ядрами мозжечка, расположенных в его черве. От ядер Голля и Бурдаха (ядра в продолговатом мозге, отвечающие за глубокую чувствительность) некоторая часть аксонов проходит также в червь мозжечка.
От нижних олив через нижние мозжечковые ножки проходят к коре мозжечка проводники экстрапирамидной системы.
Из различных отделов коры головного мозга начинаются корково-мосто-мозжечковые тракты, берущие свое начало в лобной доле (передние отделы верхней и средней лобных извилин), затылочной и височной долях (средняя и нижняя височные извилины и затылочная кора) переключаются на вторые нейроны в ядрах моста своей стороны. Аксоны вторых нейронов совершают перекрест в основании моста и через средние ножки мозжечка входят в противоположное полушарие мозжечка, где и заканчиваются в его коре.
Таким образом, полушария большого мозга оказываются связанными с противоположными полушариями мозжечка. Мозжечковые расстройства, возникающие при поражении коры (главным образом лобных и височных долей), обнаруживаются на противоположной очагу стороне.
Пути от мозжечка
Нижние и средние ножки мозжечка состоят, в основном, из проводников, направляющихся к мозжечку; верхние же ножки являются путем, по которому отходят от мозжечка его эфферентные волокна. Главными проводниками верхних ножек являются следующие.
Пути от зубчатого ядра мозжечка, совершающие перекрест и заканчивающиеся в противоположных красных ядрах и частично направляющиеся далее к зрительному бугру и далее в бледный шар и полосатое тело. Данными связями мозжечок связывается с структурами экстрапирамидной системы. От красных ядер начинается рубро-спинальный тракт (тракт Монакова), идущий от клеток красного ядра и нисходящий по мозговому стволу и спинному мозгу. Располагаясь в его боковом столбе пучок Монакова заканчивает свои волокна в передних рогах спинного мозга, устанавливая контакт с клетками периферических двигательных нейронов и, следовательно, с мускулатурой. Это — основной церебелло-спинальный путь, устанавливающий связи мозжечка с мускулатурой.
Помимо него мозжечок оказывает свое нисходящее влияние на спинной мозг опосредованно через вестибулоспинальный тракт и задний продольный пучок. Благодаря последнему мозжечок функционально связан с сочетанным движением глазных яблок и головы.
Анализируя перечисленные проводящие пути мозжечка с их перекрестами, мы можем убедиться в следующем.
Каждое полушарие мозжечка получает импульсы из одноименной половины тела
Каждое полушарие мозжечка получает импульсы от коры противоположных долей больших полушарий
Каждое полушарие мозжечка направляет импульсы к передним рогам спинного мозга и следовательно мышцам одноименной половины тела
Кроме того, в мозжечке существует соматотопическая проекция. В черве представлена мускулатура туловища, в полушариях — конечностей, особенно их дистальных отделов. Следовательно червь с его связями ответственен за функции равновесия, а полушария мозжечка -за координацию и точность движений конечностей.
Функцией мозжечка является рефлекторное поддержание мышечного тонуса, равновесия, координации и синергии движений. Поражения мозжечка или его связей сопровождаются нарушением статики – способности поддержания стабильного положения центра тяжести, обеспечивающего устойчивость и ра с строй ст вами к оординации движений
Строение спинного мозга
Спинной мозг расположен внутри позвоночного канала. Верхняя граница его (с продолговатым мозгом) соответствует уровню перекреста пирамид или месту выхода I пары шейных корешков. Нижний конец спинного мозга находится на границе I и II поясничных позвонков.
Спинной мозг представляет собой длинный тяж (длиной 42 — 45 см), окруженный тремя оболочками: твердой, паутинной и мягкой; он фиксирован внутри позвоночного канала своими корешками и зубчатой связкой.
Между паутинной и мягкой мозговыми оболочками в так называемом подпаутинном, или субарахноидальном, пространстве циркулирует цереброспинальная жидкость.
Спинной мозг состоит из 31 — 32 сегмента; каждому из них соответствуют две пары корешков — передних и задних. Различают следующие отделы или части спинного мозга:
pars cervicalis (шейная часть) — из 8 шейных сегментов (С1-C8);
pars thoracalis (грудная часть) — из 12 грудных сегментов (Th1- Th12);
pars lumbalis (поясничная часть) — из 5 поясничных сегментов (L1- L5);
pars sacralis (крестцовая часть) — из 5 крестцовых сегментов (S1- S5).
Наконец, самым нижним сегментом является один (иногда два) копчиковый сегмент (Co1-2).
Средняя величина диаметра поперечного сечения спинного мозга равняется 1 см; в двух местах этот диаметр увеличивается, что соответствует так называемым утолщениям спинного мозга.
В состав шейного утолщения входят V, VI, VII, VIII шейные и I — II грудные сегменты;
в состав поясничного — все поясничные и I — II верхние крестцовые сегменты;
три нижние крестцовые (III — V) и копчиковый сегменты составляют так называемый conus medullaris — конически суживающийся нижний конец спинного мозга.
Из спинного мозга выходит, соответственно числу сегментов, 31 пара передних двигательных корешков и входит в него 31 пара задних чувствительных корешков. Передние и задние корешки внутри позвоночного канала сближаются и собираются в общий пучок после межпозвоночного ганглия (ganglion spinale intervertebrale), расположенного в межпозвоночном отверстии. Общий пучок двигательных и чувствительных волокон из обоих корешков, выходящий из межпозвоночного отверстия, называется корешковым нервом.
В процессе роста спинной мозг отстает в длине от позвоночника и у взрослого человека оказывается много короче последнего. Нижний отдел спинного мозга оказывается расположенным на границе I и II поясничных позвонков. Соответственно этому, корешки, направляющиеся к своим межпозвоночным отверстиям, лишь в верхних отделах (шейных) расположены горизонтально. Уже начиная с грудного отдела, они идут косо книзу, а ниже conus medullaris корешки поясничных и крестцовых сегментов располагаются внутри позвоночного канала почти отвесно, составляя так называемый конский хвост (cauda equina).
Восходящие пути
Пучки Голля и Бурдаха располагаются в задних столбах, проводят импульсы глубокой чувствительности, т.е ответственны за суставно –мышечное чувство, вибрационную чувствительность, чувство веса и давления;
Спино-таламический тракт (tractus spino-thalamicus) располагается в боковых столбах, является основным проводником поверхностной чувствительности (болевой, температурной, тактильной);
Пучки Флексига и Говерса (tractus spino-cerebellaris) проходят в боковых столбах и являются афферентными проводящими путями для передачи нервного импульса от рецепторов опорно-двигательного аппарата в мозжечок, т.е. контролируют функции равновесия и координации движений.
Нисходящие пути
Основной боковой пирамидный пучок (tractus cortico-spinalis lateralis) проходит в боковых столбах спинного мозга и передает нервные импульсы произвольного движения;
Монаковский, или рубро-спинальный пучок (tractus rubro-spinalis) также расположен в боковых столбах. Ответственен за нисходящее влияние мозжечка и подкорковых ганглиев на структуры спинного мозга, а через них на мышечный аппарат, тем самым регулируя мышечный тонус, функции координации движений и равновесия;
Вестибуло-спинальный пучок (tractus vestibulo-spinalis) располагается в передне-боковых столбах, проводит нервное возбуждение от вестибулярного анализатора и мозжечка к сегментарному аппарату спинного мозга и далее к скелетным мышцам, за счет чего регулируется равновесие, координация движений;
Задний продольный пучок (fasciculus longitudinalis posterior) проходит в передних столбах спинного мозга, регулирует нисходящие связи глазодвигательных нервов, мозжечка, вестибулярного анализатора и спинного мозга, отвечает за содружественные повороты глазных яблок и головы, участвует в механизме возникновения нистагма.
Волокна всех нисходящих проводников заканчиваются у клеток передних рогов спинного мозга. Таким образом, периферический двигательный нейрон расположенный в передних рогах спинного мозга получает импульсы от вышележащих отделов нервной системы, имеющих отношение к мышечному тонусу и движению. Импульсы «произвольного» движения поступают из коры головного мозга по пирамидному пути, от экстрапирамидной двигательной системы по рубро-спинальному пучку, от мозжечка по вестибуло-спинальному и тому же рубро-спинальному пучку.
Возникающие при поражении проводникового аппарата спинного мозга расстройства носят, в отличие от сегментарных расстройств, более диффузный характер с выпадением функций нижележащих отделов нервной системы. При поражении двигательных (пирамидных) путей в состоянии центрального паралича оказываются отделы мускулатуры, иннервируемые от всех нижележащих сегментов. При перерыве чувствительных путей анестезированными оказываются участки, соответствующие также всем расположенным ниже сегментам (так называемые проводниковые расстройства чувствительности).
Поражение заднего столба (пучки Голля и Бурдаха) вызывает утрату суставно-мышечного чувства, вибрационной и тактильной чувствительности книзу от уровня поражения на стороне очага.
Поражение бокового столба спинного мозга (пирамидный и спиноталамический тракты) дает книзу от уровня поражения центральный паралич на своей стороне и утрату болевой и температурной чувствительности на противоположной.
Половинное поражение поперечника спинного мозга создает картину броун-секаровского паралича. Проводниковые расстройства выражаются в центральном параличе и утрате суставно-мышечного и вибрационного чувства на стороне поражения ниже пораженных сегментов; на противоположной очагу стороне также ниже пораженных сегментов наблюдается выпадение болевой и температурной чувствительности. Соответственно пораженным сегментам (серого вещества) на стороне очага могут наблюдаться также сегментарные двигательные (периферические параличи в соответствующих миотомах) и чувствительные расстройства в соответствующих дерматомах.
Полное поражение поперечника спинного мозга вызывает параплегию нижних конечностей или (при высоких поражениях) тетраплегию, т. е. паралич четырех конечностей с утратой всех видов чувствительности книзу от уровня поражения; одновременно возникают нарушения функции тазовых органов (мочеиспускания, дефекации). На уровне пораженных сегментов возникают сегментарные нарушения по миотомам и дерматомам, книзу от очага — проводниковые.
Функции нервной системы
1. Регуляция деятельности всех органов и систем, эндокринная и иммунная регуляция, поддержание внутренней среды организма
2. Взаимодействие организма с внешней средой, анализ различных видов внешних раздражителей
3. Ответная реакция организма на различные виды воздействий, направленная на адаптацию к окружающей среде
4. Контроль за положением тела в пространстве, регуляция всех видов двигательных актов.
5. Осуществление всех видов психической деятельности (сознание, эмоциональное реагирование, речь, память, гнозис, праксис, мышление, интеллект, особенности человеческой личности).
Нервная клетка
Нервную ткань составляют нервные клетки (нейроны) с их отростками и клетки глии. Сосуды и оболочки мозга имеют мезодермальное происхождение.
Нейрон является основной структурной и функциональной единицей нервной системы и состоит из тела (сома) и отростков. Как правило имеются один длинный маловетвящийся центральный отросток –аксон и короткие периферические, имеющие много ответвлений дендриты.
Только в головном мозге насчитывается около 10 млрд нейронов.
Основные функции нейрона:
-проведение нервного импульса
-синтез белков, липидов, углеводов
-синтез нейромедиаторов в некоторых случаях гормонов.
Нейроны делятся на сенсорные (чувствительные), моторные (двигательные) и ассоциативные, а по характеру вырабатываемого в них медиатора дифференцируются на холинергические, серотонинергические, адреналинергические, ГАМКергические, допаминергические и.т.п.
Тело клетки и ее отростки покрывает непрерывная сдвоенная мембрана (невролемма), выполняющая разграничительную и транспортную функцию. Благодаря мембране осуществляется пассивный транспорт воды и низкомолекулярных веществ, а также работа ионных насосов для поддержания внутри нейрона постоянного мембранного потенциала покоя и возникновения возбуждающего потенциала действия с резким изменением проницаемости для содержащих биоэлектрический заряд ионов, что определяет возникновение нервного импульса.
В теле нейрона располагается цитоплазма и ядро с ядрышком, содержащие ДНК и РНК. В ядре сконцентрирован генетически аппарат клетки.
Комплекс Гольджи участвует в синтезе углеводных полимеров и гормонов. Митохондрии содержат дыхательные ферменты, обеспечивают процесс окислительного фосфорилирования и участия в гликолизе, их основная функция синтез богатой энергией АТФ. Лизосомы, содержащие гидролитические ферменты и расщепляют для последующей биоутилизации биологические макромолекулы внутриклеточного и внеклеточного происхождения. Эндоплазматическая сеть с гранулами РНК и рибосомами ответственны за синтез белков.
Нервное волокно-в большинстве случаев аксон нервной клетки, состоящий из осевого цилиндра, покрытого миелиновой оболочкой. Миелиновая оболочка состоит из спиралевидно окружающих осевой цилиндр аксона слоев, при этом толщина аксона, в основном, зависит от толщины миелиновой оболочки и составляет от 0,3 до 20 мкм. Миелиновая оболочка аксона продуцируется из оболочек прилегающих к нейрону глиальных клеток, в центральной нервной системе –олигодендроцитов, в периферической нервной системе-шванновских клеток (леммоцитов). Между фрагментами миелиновой оболочки образуются просветы, лишенные миелина-перехваты Ранвье. Скорость проведения по волокну нервного импульса прямо пропорциональна толщине его миелиновой оболочки и варьирует от 0,6 до 20 м/с. Наибольшей скоростью проведения обладают толстые миелинизированные волокна. Тонкие слабомиелинизованные нервные волокна проводят импульс с существенно меньшей скоростью.
Для нейрона характерна исключительно высокая активность метаболических процессов. Биосинтез большинства макромолекул осуществляется в теле нервной клетки. Отсюда они распространяются по аксону посредством ортоградного аксонального тока (аксональный транспорт). Благодаря аксотоку по аксону транспортируются молекулы белка, гликопротеиды, фосфолипиды, некоторые ферменты, необходимые для поддержания метаболизма. Продукты, происходящих в аксоне процессов метаболизма посредством ретроградного аксонального тока перемещаются в тело клетки, где подвергаются дальнейшему разрушению ферментами лизосом до составных эелементов, используемых при формировании вновь синтезируемых в теле нейрона макромолекул.
Нервная система способна преобразовывать воздействие внешних и внутренних раздражающих факторов в энергию нервного импульса, имеющую электрическую природу.
При этом нервный импульс перемещается всегда в одном направлении: от дендритов к телу клетки, а затем по аксону.
Нервный импульс и синаптическая передача
Превращение различных видов энергии в биоэлектрическую энергию происходит в рецепторах, являющихся по сути биологическими преобразователями. В рецепторах потенциал действия возникает если первоначальный раздражитель превысил пороговую величину.
Нервный импульс генерируется и передается в форме потенциала действия. Было установлено, что разность потенциалов на наружной и внутренней стороне мембраны нервного волокна, определяющая его электрический заряд, поддерживается постоянно и зависит от происходящих в мембране активных химических процессов, обеспечивающих перемещение ионов через мембрану в направлении, противоположном градиентам их концентрации. При этом мембрана проницаема не для всех ионов в равной степени. Так, К мембрана пропускает легче, чем Na, Ca. Вместе с тем внеклеточная жидкость содержит много ионов калия и меньше ионов натрия. В итоге в периоды покоя в клетке преобладают отрицательно заряженные ионы хлора, фосфатов и органических кислот, в результате потенциал покоя внутри клетки отрицательный. Под влиянием медиатора в синаптической щели и активизации рецепторов постсинаптической мембраны происходит прорыв внутрь нервной клетки положительно заряженных ионов, главным образом Na, что ведет к изменению отрицательного внутриклеточного электрического заряда на положительный. Это явление называют деполяризацией клеточной мембраны, возникает потенциал действия, что является физической основой возбуждения.
При деполяризации мембраны в зонах перехватов Ранвье происходит «подзарядка» нервного импульса. В результате возникает сальтаторное (скачкообразное) перемещение импульса без декремента, т.е. без потери энергии. Другими словами, нервный импульс перемещается по нервному волокну, не затухая, без потери энергии.
Место, где окончание аксона сближается с дендритом или телом следующего нейрона называется синапсом. Синапсы обеспечивают усиление или торможение потока нервных импульсов и определяют проведение их всегда в одном направлении.
Аксон заканчивается утолщением (пресинаптическая пуговка), в котором содержатся митохондрии и пузырьки с квантами медиатора. Участок пресинаптической пуговки максимально приближенный к невролемме следующего нейрона называется пресинаптической мембраной. Между пресинаптической и постсинаптической мембранами имеется узкая синаптическая щель. Когда нервный импульс доходит до пресинаптической пуговки из расположенных в ней синаптических пузырьков в синаптическую щель выделяется квант нейромедиатора, который достигает постсинаптической мембраны и меняет ее проницаемость для находящихся вокруг положительно и отрицательно заряженных ионов, вызывая в расположенном по другую сторону синаптической щели нейроне возникновение возбуждающего или тормозного постсинаптического потенциала. Таким образом, нейромедиатор обеспечивает химическую передачу нервного импульса через синаптическую щель. По влиянию на синаптические аппараты нейромедиаторы бывают возбуждающими (глутамат, аспартат) и тормозными (ГАМК, глицин). Другие нейротрансмиттеры, состоящие главным образом из моноаминов (ацетилхолин, дофамин, норадреналин, серотонин) могут, воздействуя на одни нейроны, оказывать возбуждающее действие, тогда как влияние их на другие нейроны может быть тормозным.
Таким образом, влияние нейромедиатора на постсинаптическую мембрану может обусловить не только ее деполяризацию в связи с прохождением положительно заряженных ионов, но и нарастание отрицательного биопотенциала в тех случаях, когда, через нее в клетку проникают в основном отрицательно заряженные ионы. В таких случаях деполяризации мембраны не происходит, и клетка сохраняет, и даже наращивает свой потенциал покоя, т.е. нейрон тормозится, становится менее возбудимым.
Помимо нейротрансмиттеров на синаптическую передачу могут оказывать усиливающее или ослабляющее действие нейромодуляторы (эндорфины, соматостатин, субстанция Р) и нейрогормоны (ангиотензин, вазопрессин).
Необходимо выделить два типа нервной передачи-импульсный и безыимпульсный. Импульсная передача обеспечивается электролитным и нейромедиаторным механизмами, безымпульсная осуществляется посредством аксонального транспорта, функция которого преимущественно трофическая.
Нейроглия
Глиальных клеток в нервной системе примерно в 10 раз больше, они не имеют аксонов и сохраняют способность к делению в постнатальном периоде. Выделяют астроглию, олигодендроглию и эпендимную глию.
Астроглия состоит из астроцитов, которые участвуя в формировании гематоэнцефалического барьера (ГЭБ), выполняют важную роль в обеспечении метаболических процессов в нервных клетках. Астроциты располагаясь между нейронами и капиллярами, избирательно пропускают в нейрон биологически активные вещества и продукты обмена. Было доказано участие астроцитов в метаболизме глутамата и ГАМК, которые являются соответственно возбуждающими и тормозными медиаторами. При нарушении иммунных процессов в нервной ткани астроциты приобретают свойства иммуноэффекторных клеток, экспрессирующих ряд антигенов и иммуномодуляторов, которые формируют в мозговой ткани иммунный ответ, что играет роль в патогенезе аутоиммунных демиелинизирующих заболеваний нервной системы.
Олигодендроглия сконцентрирована в белом веществе мозга, формирует миелиновую оболочку нервных волокон. На периферии функции олигодендроцитов выполняют шванновские клетки (леммоциты).
Эпендимная глия выстилает расположенные внутри мозга ликворные пути (желудочки и водопровод мозга, а также центральный канал спинного мозга) и выполняет, таким образом, барьерную функцию.
Глия выполняет, главным образом, опорную функцию, формируя своеобразный каркас, в ячейках которого находятся тела нейронов, обеспечивает относительную изоляцию и обособление нейронов, поддержание ионного баланса, выполняет аккумулирующую функцию, поглощая выделяемые нейронами при их «зарядке» и «разрядке» свободные электролиты. Глиальные элементы, взаимодействуя с нейронами, обеспечивает транспортные функции, обладают способностью к накоплению РНК, белка, дыхательных ферментов, обеспечивают возможность поглощения и накопления запасов медиаторов с последующим их высвобождением. Глия (в основном леммоциты) участвует в процессах дегенерации и регенерации нервной ткани. Есть мнение о том, что глия является местом хранения закодированной информации, что обеспечивает, так называемую долговременную память.
Разрастание глиальной ткани при повреждении мозга ведет к формированию мозгового рубца. Установлено также, что астроциты участвуют в поддержании ионного баланса и, следовательно, влияют на генерацию нервного импульса, регулируя уровни Na+ и K+ вокруг клетки. Считается, что формирование глиальных рубцов нарушает забуферивание K+ вокруг нервной клетки, что приводит к снижению порога возбудимости нейрона и эпилептическим разрядам.
Таким образом, изменение состояния и функции глии может влиять на развитие многих физиологических процессов и заболеваний нервной системы.
Гематоэнцефалический барьер
Эндотелий капилляров и расположенных в мозговых желудочках сосудистых сплетений вместе с элементами глиальной ткани (главным образом астроцитами) составляют гематоэнцефалический барьер (ГЭБ).
Контакты между эндотелиальными клетками достаточно плотные, промежутки между ними, как правило, меньше 1,5 нм, что служит препятствием для большинства молекул, в частности белковых. К тому же между капилляром и нейроном располагается астроцитарная глия. Эти структуры служат своеобразным фильтром, обеспечивающим относительную защиту нервных клеток и их отростков.
Таким образом, ГЭБ регулирует поступление в цереброспинальную жидкость и нервную ткань циркулирующих в крови веществ, обеспечивая относительное постоянство внутренней среды мозга. ГЭБ защищает мозг и цереброспинальную жидкость от всевозможных случайных эндогенных и экзогенных факторов, которые, попав в кровяное русло, могли бы оказать на нервную систему неблагоприятное воздействие.
При патологических состояниях проницаемость ГЭБ может увеличиваться, что обусловливает проникновение в нервную систему веществ, приводящих к нарушению в ней гомеостаза и развитию различных патологических состояний (отек, набухание, аутоиммунные процессы и др.). Проницаемость ГЭБ играет роль и в лечении патологических процессов нервной системы, т.к. для достижения терапевтического эффекта необходимо учитывать возможность проникновения того или иного лекарственного средства через ГЭБ.
Строение нервной системы
Нервная система анатомически состоит из центрального и периферического отделов. Кроме того выделяют соматическую (анимальную) и вегетативную (автономную) нервную системы. Соматическая нервная система регулирует произвольные движения поперечно-полосатой мускулатуры скелета, языка, гортани, глотки, обеспечивает общую чувствительность и связь с внешней средой. Вегетативная нервная система в большей степени влияет на функции гладких мышц внутренних органов, сосудов, сердечную деятельность, секреторную функцию железистой ткани, в том числе и эндокринных желез, регулирует обменные процессы во всех органах и тканях. Вегетативная нервная система не подчиняется воле человека, а потому считается автономной и подразделяется на симпатический и парасимпатический отделы, действующие функционально антагонистически.
Центральная нервная система это расположенный в полости черепа головной мозг и находящийся в позвоночном канале спинной мозг.
К периферической нервной системе относятся парные спинномозговые и черепные нервы с их корешками, их ветви, образующие нервные сплетения и периферические нервы, нервные окончания и ганглии. Периферические нервы в зависимости от того, из каких волокон (чувствительных либо двигательных) они состоят, подразделяются на чувствительные, двигательные и смешанные. Тела нейронов, отростки которых составляют чувствительные нервы, лежат в нервных узлах вне мозга. Тела двигательных нейронов лежат в передних рогах спинного мозга или двигательных ядрах головного мозга.
Макроскопически головной и спинной мозг состоят из серого и белого вещества. Серое вещество представляет из себя скопление тел нервных клеток. Отдельные ограниченные скопления серого вещества в головном мозге носят названия ядер, в спинном мозге соответственно называются рогами.
Поверхность головного мозга –серая, благодаря наличию множества тел нейронов. К серому веществу головного мозга относят кору больших полушарий, базальные (подкорковые) ганглии, ядра зрительных бугров и гипоталамуса, скопления тел нейронов в стволе головного мозга (ядра черепных нервов, ретикулярная формация, бугорки четверохолмия, красные ядра, черная субстанция). Серое вещество спинного мозга представлено скоплениями тел нейронов центральной части сегмента спинного мозга и локализовано в передних, боковых и задних рогах спинного мозга.
Белое вещество-это отростки нервных клеток (аксоны) с миелиновыми оболочками, составляющие проводящие пути головного и спинного мозга.
Дата: 2019-04-23, просмотров: 349.