Описано множество генетических вариантов молекул гемоглобина с неизмененным сродством к кислороду. Фетальиый гемоглобин (HbF), однако, является примечательным исключением.
Гемоглобин F имеет две гамма (у)-цепи, а не (3-цепи, характерные для НЬА. Кривая диссоциации оксигемоглобина для HbF сдвинута влево. Возможно, это происходит из-за того, что 2,3-ДФГ не способен связаться с у-цепями, что обеспечивает высокое сродство гемоглобина к О2. Данное обстоятельство имеет существенное значение для плода, когда Рас )2 низкое и плацентарное поглощение О2 возрастает за счет высокого сродства к О2.
Доставка кислорода к тканям и его утилизация
До сих пор обсуждение касалось факторов, определяющих содержание кислорода, т. е. количество О2, связанного с гемоглобином и растворенного в плазме. Для обеспечения обмена веществ в периферических тканях ключевой физиологической переменной является доставка кислорода: количество О2, транспортируемое к тканям в единицу времени. Чтобы поддержать аэробный метаболизм и предотвратить образование молочной кислоты в анаэробном цикле, периферические ткани должны постоянно снабжаться О2. Обстоятельства, которые определяют адекватность О2-снабжения, разнообразны и включают состояние покоя, физическую нагрузку, ги-перкатаболические состояния и инфекцию.
Системная доставка кислорода
Доставка кислорода к периферическим тканям зависит от количества О2 в определенном объеме крбви и уровня кровотока. Хотя объем кровотока к отдельным органам различен, в периферических тканях он в целом равен минутному сердечному выбросу (МСВ). Системная доставка кислорода (D()2) рассчитывается как:
Do2 = МСВ (л/мин) х О2 содержание (мл/л). [ 10-7]
Общее содержание О2 равняется сумме О2, связанного с гемоглобином, и О2? растворенного в плазме. Следовательно, уравнение [10-7] может быть представлено
как:
Do? = МСВ х ([(НЬ) х 1.34 х % насыщения] + [0.0031 х Рао2]). [ 10-8]
Утилизация кислорода тканями и его артерио-венозмая разница
В условиях основного обмена взрослый человек потребляет около 250 мл О2 в 1 мин. Однако скорости утилизации О2 различными тканями значительно отличаются.
В зависимости от состояния организма (покой, нагрузка или заболевание) меняется фракционное рапределение сердечного выброса к органам. Более того, и эк-(гпракция кцслорода тканями различных органов неодинакова. Например, миокард получает лишь малую фракцию сердечного выброса, но извлекает почти весь доставляемый О2. Измерение содержания О, в венозной крови, покидающей миокард (н коронарном синусе), выявило большую артерио- венозную разницу кислорода.
Без помещения катетера непосредственно в вену, отходящую от того или иного < 'ргана, нельзя рассчитать органную артерио-венозную разницу О> и, следовательно,
катетера правых отделов сердца позволяет получать пробу крови из правого желудочка или легочной артерии и измерять содержание кислорода или насыщение смешанной венозной крови (гл. 12). Содержание кислорода в смешанной венозной крови представляет собой усредненную величину для венозной крови от всех органов — и с низким, и с высоким уровнями экстракции О2.
Правило Фика
Существует тонкое сопряжение между артериальным содержанием О2, сердечным выбросом, тканевой утилизацией О2 и содержанием О2 в смешанной венозной крови. Некоторые заболевания, такие как респираторный дистресс-синдром взрослых (гл. 14) и сепсис, сопровождаются нарушением сопряжения между утилизацией О2 периферическими тканями и доставкой кислорода. Утилизация снижается, когда доставка падает ниже некоторого порога.
Отношение между этими переменными выражается правилом Фика, которое устанавливает, что потребление О2 (объем в 1 мин) является произведением минутного сердечного выброса и артерио-венозной разницы О2:
Vo. =Qx(Cao2 -Cvo2), [10-9]
где: СаО2 — содержание О2 в артериальной крови (мл О2/л),
Cvo2 — содержание О2 в смешанной венозной крови (мл О2/л), Q ~ минутный сердечный выброс (МСВ).
Анализ уравнения Фика показывает, что возросшая кислородная потребность при фиксированном минутном сердечном выбросе вызывает увеличение артерио-венозной разницы О2. Напротив, снижение метаболических требований при фиксированном минутном сердечном выбросе влечет за собой уменьшение этой разницы. Отсюда содержание О2 в смешанной венозной крови и, следовательно, ее насыщение кислородом и РО2 зависят от экстракции кислорода тканями и доставки к ним О2. Используя уравнение [10-9], рассчитаем артерио-венозную разницу кислорода для здорового человека с минутным сердечным выбросом 5 л и потреблением О2 250 мл/мин:
250 мл/мин = 5 л/минх (Сао2- Cvo2), [10-10]
(Сао2 - Cvo2) = 50 мл О2/л = 5 мл/дл. [10-11]
Если содержание гемоглобина 150 г/л и насыщение артериальной крови кислородом нормально, то СаО2 составит 200 мл/л. Решая уравнение [10-11] относитель-но CvO.,, получаем величину 150 мл/л. С помощью кривой диссоциации оксиге моглобина (для венозной крови) определяем соответствующее РО2 смешанной ве нозной крови (PvO2) — 40 мм рт. ст.
В клинической практике содержание О2 в смешанной венозной крови измеря ют, катетеризируя правые отделы сердца. Содержание О2 в артериальной крови он ределяют на основании анализа газов артериальной крови. Используя уравнена Фика, рассчитывают МСВ. Скорость потребления О2 принимается равно) 250 мл/мин. Потребление О2 может быть рассчитано по результатам анализа кон центрации О2 в выдыхаемом воздухе. Катетеризация легочной артерии также ис пользуется в клинике для расчета МСВ методом термодилтоции (гл. 12).
Транспорт двуокиси углерода
[рчнячительное количество СО2. Следовательно,
разделе рассматриваются транспорт СО2, включая многочисленные формы СО2 в крови, и кривая диссоциации СО2.
Транспортные формы СО2 крови
Двуокись углерода легко диффундирует. Ее способность к диффузии в 20 раз превышает таковую у кислорода. По мере образования в процессе клеточного метаболизма СО2 диффундирует в капилляры и транспортируется к легким в трех основных формах: (1) в виде растворенной СО2, (2) в виде аниона бикарбоната и (3) в виде карбаминовых соединений (рис. 10-4).
Растворенная СО2
СО2 очень хорошо растворяется в плазме. Количество растворенной в плазме СО2 определяется произведением ее парциального давления и коэффициента растворимости (а = 0.3 мл/л крови/мм рт. ст.). Около 5 % общей двуокиси углерода в артериальной крови находится в форме растворенного газа.
Анион бикарбоната
Эта форма СО2 является преобладающей (90 %) в артериальной крови. Бикар-бонатный анион является продуктом реакции СО2 с водой с образованием Н2С(>{ и ее диссоциации на водород и ион бикарбоната:
СО2 + Н2О «~> Н2СО3о Н+ + НСО3". [10-12]
Реакция между СО2и Н2О протекает медленно в плазме и очень быстро в эритроцитах, где присутствует внутриклеточный фермент карбоашидраза. Она облегчает реакцию между СО2 и Н2О с образованием Н2СОЛ; вторая фаза уравнения [ 10 -121 протекает быстро без катализатора.
По мере накопления HCO:i внутри эритроцита анион диффундирует через клеточную мембрану в плазму. Мембрана эритроцита относительно непроницаема для ЬГ, как и вообще для катионов, поэтому ионы водорода остаются внутри клетки. Электрическая нейтральность клетки в процессе диффузии НСО3 в плазму обеспечивается притоком ионов хлора из плазмы в эритроцит, что формирует так называемый хлорид ный сдвиг (рис. 10-4).
Часть FT, остающихся в эритроцитах, забуферивается, соединяясь с гемоглобином. В периферических тканях, где концентрация СО2 высока и значительные количества Нг накапливаются эритроцитами, связывание Н1 облегчается деоксигенаци-ей гемоглобина. Восстановленный гемоглобин лучше связывается с протонами, чем оксигенированный. Таким образом, деоксигенация артериальной крови в периферических тканях способствует связыванию Hf посредством образования восстановленного гемоглобина. Это увеличение связывания СО2 с гемоглобином известно как эффект Холдейна (рис. 10-5).
Карбаминовые соединения
Третьей формой транспорта СО2являются карбаминовые соединения, образованные в реакции СО2 с концевыми аминогруппами белков крови. Основным белком крови, связывающим СО2> является гемоглобин (особенно глобимовая часть молекулы). Этот процесс описывается следующей реакцией:
Hb-NH2 + СО 2 <-> Hb-NH COOH <-> Hb-NHCOO + Н4 [10-13]
Реакция СО;,с аминогруппами иротекаетбысфо. Как и вслучае бо.чге лп кого
160 _____
Рис. 10-4. С02-
транспорт в крови, иллюстрирующий образование НСО~ и кар-баминовых соединений, хло-ридиый сдвиг и святил ванне Н + . При hoi-лощении О2 и высвобождении СО2 и легочных капиллярах реакции, представленные па схеме, протекают в обратном па-правлении
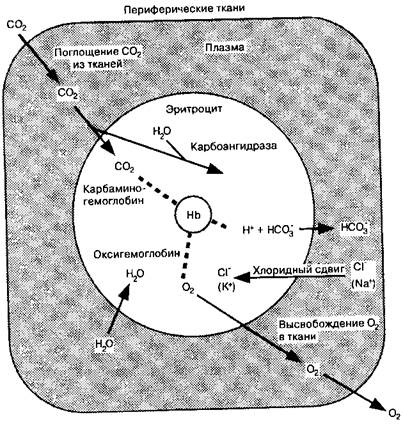
единений легче протекает с деоксигенированными формами гемоглобина. Карбами-новые соединения составляют около 5 % общего количества СО2, транспортируемого артериальной кровью.
С точки зрения сравнительного вклада каждой из этих форм в артерио-венозную разницу по концентрации СО2, около 60 % принадлежит HCCV, карбаминовым соединениям - около 30 % и растворенной СО2 - около 10 %.
Кривая диссоциации СО2-гемоглобин
Присутствие в крови всех трех форм СО2 создает равновесие между растворенной СО2 (РСО2) и СО2, химически связанной с другими веществами (рис. 10-5).
В отличие от S-образной кривой диссоциации оксигемоглобина, кривая диссоциации СО2-гемоглобина более линейна. Особое физиологическое значение имеет то, что общее содержание СО2 при любом уровне РСО2 зависит от степени оксигена-ции гемоглобина (эффект Холдейна).
По мере того, как в периферических тканях происходит отщепление О2 от гемоглобина, он присоединяет СО2все более активно. Отсюда, общее содержание СО2в крови больше при любом уровне тканевого РСО2. В этом можно убедиться, сравнивая кривые диссоциации СО? для полностью деоксигенированной, полностью окси-генироианной и частично оксигенированной крови с РО2, эквивалентным смешанной венозной крови (рис. 10-6).
о ~.,,..„„ч,т,1,^пг»1;и.1гп.т аптрпмя лмтй кпови с РаСО?40 мм рт. ст. содержание СО^
Рис. 10-5. Эффект Холдейна. Связывание СО2 с гемоглобином увеличивается при деоксигепации артериальной крови. Содержание СО2 выше при данном Рассь в деоксигенированной крови
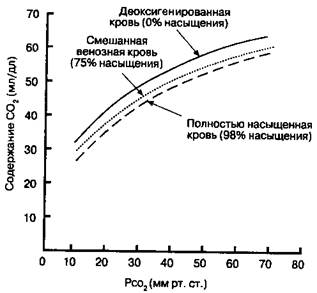
ферических тканях, кровь, прошедшая капилляры, насыщена кислородом на 75 %. При РСО2 около 46 мм рт. ст. содержание СО2 в смешанной венозной крови (т. е. крови, входящей в правый желудочек и представляющей смесь венозной крови от всех тканей тела) составляет приблизительно 530 мл/л (рис. 10-6, точка Б). Повышенное сродство деоксигенированной крови к СО2 объясняет добавочные 20-30 мл СО2/л сверх того, что могло бы быть поглощено без изменения насыщения оксигемоглобина.
О2-СО2 диаграмма
Соотношения между связыванием и транспортом О2 и СО2 обобщает О2-СО2 диаграмма, представляющая собой график содержания О2 и СО2 в крови в зависимости от их парциальных давлений (рис. 10-7).
В качестве примера рассмотрим воздействие изменений в уровне РаСО2 па содержание О2 при РаО2 40 мм рт. ст. При выраженной гипокапнии (РаСО210 мм рт. ст.) и при РаО2в 40 мм рт. ст. содержание О2 в крови составляет около 180 мл/л. При РаСО2 40 мм рт. ст. та же самая величина Ро2 обеспечивает содержание О2 только 150 мл/л (эффект Бора).
Теперь рассмотрим воздействие изменений в уровне РаО2 на содержание в крови СО2 при различных уровнях РаСО2. При РаСО2 40 мм рт. ст. и РаО, 80 мм рт. ст. (как в артериальной крови) содержание СО2 составит 500 мл/л. Когда же РаО2 равно 40 мм рт. ст., а РаСО2 — 46 мм рт. ст. (как в смешанной венозной крови), то содержание СО2 приблизится к 540 мл/л (эффект Холдейна).
Дата: 2019-03-05, просмотров: 368.