(главным образом хвостатого ядра). На ЭЭГ наблюдается усиление бета-активности (преимущественно β1) появление острых волн и вспышек тета- и дельта-волн (преимущественно в лобных отделах мозга). Электрическое раздражение стрио-паллидарных структур частыми стимулами вызывает десинхронизацию ритмов ЭЭГ. Раздражение редкими стимулами - появление вспышек веретен медленных волн. Фармакологически вызванное угнетение функций хвостатого ядра приводит сначала к появлению на ЭЭГ вспышек волн с частотой 20
54
или 14 в сек., а затем диффузное доминирование колебаний β1.
При поражениях в области зрительного бугра и III-го желудочка (с наличием клинически выраженного диэнцефального синдрома). На ЭЭГ, как правило, имеют место: пространственная, амплитудная и чаcтотная дезорганизация альфа-активности, диффузное усиление тета и дельта-активности, особенно в лобных отделах мозга, появление разнообразных форм пароксимальной активности; формирование «веретен» в лобных отделах мозга.
При поражениях в области гипоталамуса. При клинически выраженных симптомах поражения заднего гипоталамуса (сосудистая форма гипоталамического синдрома) на ЭЭГ отмечается ослабление альфа-активности и усиление бета-активности (особенно /3,). При клинически выраженных симптомах поражения переднего гипоталамуса (эндокринная форма гипоталамического синдрома) альфа-ритм на ЭЭГ обильный, но недостаточно регулярный по частоте, его зональные различия сглажены. При двустороннем разрушении гипоталамуса у экспериментальных животных наблюдается диффузная синхронизация ритмов ЭЭГ. При одностороннем
нарушении эффект синхронизации выражен ипсилатеральна. Раздражение заднего гипоталамуса электрическими импульсами, по данным одних авторов, соответствует возбуждению симпатического раздела вегетативной нервной системы и сопровождается десинхронизацией биопотенциалов. По данным других авторов, электрическое раздражение заднего гипоталамуса приводит к усилению альфа- и бета -активности. Раздражение электрическими импульсами переднего
55
гипоталамуса соответствует возбуждению парасимпатического раздела вегетативной нервной
системы и сопровождается синхронизацией биопотенциалов.
При поражениях в области задней черепной ямки «мостомозжечкового угла» и IV желудочка.
На ЭЭГ наблюдается увеличение амплитуд альфа- и тета-активности в лобной области контралатерально очагу поражения и усиление бета-активности ипсилатеральна Кроме того, отмечаются вспышки волн в височной и теменно-затылочной областях двусторонне.
При поражениях мозжечка.
На ЭЭГ-десинхронизация биопотенциалов и при этом нередко усиление дельта-активности в теменно-затылочной области ипсилатеральна
В действительности интересующие нас взаимоотношения сложнее и разнообразнее. С одной стороны, потому, что в пределах каждой макроструктуры существует много микроструктур (например, многочисленные ядра в пределах зрительного бугра). С другой стороны, существуют взаимодействия (синергия и антагонизм) между разными структурами мозга. В силу этого содержательная трактовка каждой индивидуальной записи ЭЭГ представляет нелегкую задачу. Ибо ни один из регистрируемых на ЭЭГ отдельных ее компонентов или паттернов не имеет однозначной причины своего возникновения и не отражает функционального состояния какой-либо одной структуры мозга.
- исследованиях В. Е. Майорчик [32] эта идея
выражена следующим образом. Фоновая электроэнцефалограмма говорит о самом факте и степени вовлечения в патологический процесс стволовых структур
56
мозга. Но трудно при этом конкретизировать, какая именно структура повреждена, т . к. нет специфических признаков на ЭЭГ, характерных для поражения каждой отдельной структуры. Существуют лишь некоторые недостоверные тенденции в изменениях ЭЭГ, возникающие при той или иной локализации патологического процесса.
Итак, в повседневной клинической практике мы не только не можем на основании показателей ЭЭГ ставить диагноз заболевания, но даже о локализации патологического процесса должны судить с большей осторожностью. В результате этого создается неудовлетворенность эффективностью использования метода электроэнцефалографии.
Неудовлетворенность возникает из-за того, что при клинически твердо диагностированной патологии мозга в ЭЭГ не находят отклонений от нормы в 13-60% случаев (по данным разных авторов). Наряду с этим нарушения ЭЭГ нередко в 25-49% случаев (также по данным разных авторов) обнаруживаются среди контингентов практически здоровых людей не только пожилого, но и молодого возраста Эти данные взяты нами из обзора литературы, опубликованного Bourdillon P.Y. в 1979 году [58] под названием «Электроэнцефалография сегодня».
В в то же время литературные данные убеждают в том, что биоэлектрическая активность мозга является тонким показателем деятельности мозга, очень важным для клиники.
На основании многолетних электроэнцефалографических исследований в клинической практике; мною предлагаются следующие соображения о путях повышения эффективности использования метода
57
электроэнцефалографии.
§ 2.
Исторически, традиционно сложилось так, что электрофизиологи, при проведении ЭЭГ обследований, решают три вопроса: Соответствует ли ЭЭГ общепринятым стандартам нормы, если нет, то какова степень отклонения от нормы; обнаруживаются ли у пациента признаки грубого, очагового поражения мозга и какова топика этого поражения; говорят ли данные ЭЭГ о наличии у пациента эпилептической болезни или эписиндрома, или хотя бы «повышенной судорожной готовности мозга».
Статистика показывает, однако, что в широкой клинической практике (т. е. не в специализированных нейрохирургических, неврологических и психиатрических стационарах), пароксизмальные формы биоэлектрических потенциалов или грубые макроочаговые нарушения ЭЭГ встречаются относительно редко. Чаще (70-80% случаев) наблюдаются диффузные изменения биоэлектрической активности мозга, не носящие очагового и эпилептического характера. Что же может сказать электрофизиолог в этих случаях?
Ответ на этот вопрос может быть получен, если
отталкиваться от следующих теоретических представлений.
Среди факторов, определяющих формирование целостных паттернов ЭЭГ, следует различать местные и дистантные влияния:
а) Местные влиянии: кровообращение и лимфообращение в мозговой ткани; состав и соотношение нейрогуморов и медиаторов; циркуляция нервных импульсов по коротким нейрональным связям. Эти и
58
другие местные влияния определяют метаболизм нейронов и влияют на их мембранные потенциалы.
Местные влияния выражены как в норме, так и в патологии. В первом случае они участвуют в формировании нормальных паттернов ЭЭГ. Во втором случае важно различать макро- и микропроцессы. Макропроцессы могут быть диффузными (дегенеративные, нейроинфекционные и др.). Они же могут быть очаговыми, локальными (опухоль/инсульт и т. п.).
§ больных, не имеющих грубого, деструктивного поражения мозга, можно предполагать наличие микроструктурных нарушений на нейрональном уровне. Это может иметь место у больных с выраженным атеросклерозом, с преходящими ишемическими атаками, у больных, перенесших нейроинфекции или закрытую травму мозга. Нарушение стенок капилляров мозга или микроскопические очажки поражения мозговой ткани, безусловно, возникают у них и сопровождаются затем нейродинамическими расстройствами в виде изменения скорости, регулярности и направления движения процессов возбуждения в системах корковых нейронов. Во всех таких случаях патологии происходят сдвиги в протекании биоэлектрической активности мозга.
б) Отдаленные, дистантные влияния: весь тот поток импульсаций, который поступает в кору по афферентам, как специфическим, так и неспецифическим нервным путям и влияет на мембранные потенциалы.
Со времени первых блестящих исследований Мэгуна, Моруцци, Джаспера прошло много лет. Новейшие представления о лимбико-ретикулярном комплексе далеко выходят за границы двух первоначально описанных неспецифических систем: мезэнцефалической и
59
таламической ретикулярной формации. Лимбико-ретикулярный комплекс в настоящее время рассматривается как обширная, многозвеньевая система, включающая структуры продолговатого мозга,
понтомезэнцефалические, таламические и гипоталамические образования, некоторые структуры лимбической системы, особенно неостриатум (главным образом хвостатое ядро) и даже некоторые зоны ассоциативной коры. Все, что известно в настоящее время
морфологии и физиологии этих образований, фармакологическое их изучение, многочисленные острые и хронические опыты на животных и клинические данные, особенно в области психиатрии, все это указывает на тесную связь деятельности лимбико-ретикулярного комплекса с фоновой электроэнцефалограммой.
Характерные для взрослых паттерны ЭЭГ складываются у человека в течение 10-13 лет после его рождения. Они в значительной степени генетически обусловлены и в известной мере характеризуют тип высшей нервной деятельности и индивидуальные
особенности человека. Сложившиеся после морфофизиологического созревания паттерны ЭЭГ могут трансформироваться под действием различного рода патологических воздействий. Как морфофункциональное, физиологическое созревание корково-подкорковых и межцентральных взаимоотношений, так и перестройка деятельности коры в патологических условиях, происходит при непосредственном участии лимбико-ретикулярного комплекса.
Дистантные влияния выражены как в норме, так и а патологии. В норме они формируют нормальные паттерны,
§ нормальными компонентами.
60
патологии для некоторых категорий больных трудно предположить наличие местных, микроструктурных и даже обменных нарушений вещества мозга. Это относится, например, к больным с неврастенией и другими неврозами, больным с остеохондрозом, сирингомиелией и многими другими заболеваниями. В этих случаях возможен и другой механизм возникновения сдвигов на ЭЭГ, вследствие изменения импульсных посылок в кору со стороны нижележащих, регулирующих систем мозга. Изменения на ЭЭГ вызываются в этих случаях дистантно, со стороны различных, далеко расположенных срединных структур — базальных ядер, лимбической системы, диэнцефалона, ствола мозга, и всех других звеньев лимбико-ретикулярного комплекса. Можно думать, что такие влияния зависят от дисфункции в деятельности этих структур, связанных (как и в коре) с нарушениями кровоснабжения, и с наличием микроочаговых поражений этих систем. Диапазон изменений ЭЭГ при этом очень велик, к ним относятся: значительная дезорганизация альфа-активности по всем ее параметрам (частоте, амплитуде, форме волн, извращению зональных различий); значительное усиление бета-активности низкой частоты; наростание индекса и амплитуды тета- и дельта-волн; появление пиков, острых волн, комплексов, вспышек волн
в разных областях мозга. Изменения ЭЭГ такого генеза мы называем регуляторными.
Существуют, наконец, и разные комбинации влияний на ЭЭГ разных факторов – местного, микроструктурного и регуляторного.
Эти представления теснейшим образом связаны с концепцией о двух формах патологии, наиболее четко сформулированной в работах А. М. Зимкиной с
61
сотрудниками [34, 35]. Суть концепции в следующем. Существуют нарушения конкретных частных функций ц.н.с. (двигательных, речевых и т. д). Такие нарушения обычно наступают вследствие более или менее обширной деструкции вещества мозга. И существует изменение общего функционального состояния мозга, ц.н.с, выражающееся в дисфункции, дисбалансе в деятельности самых разных систем мозга – сердечно-сосудистой, нейро-гуморальной и т. д. Первая форма патологии является первичным , органическим ядром заболевания. Вторая формирует широкий класс « болезней регуляции», которыми ядро заболевания как бы обрастает.
Практика показывает, что первичное органическое ядро заболевания далеко не всегда играет решающую роль
§ состоянии пациентов. Для последующей деятельности не всегда важно то что утрачена Важнее – степень компенсации дефекта, степень использования резервных возможностей, которые очень велики. Болезни регуляции оказываются ведущими в поликлинической практике у больных с такими диагнозами, как астено-невротические состояния, вегето-сосудистые расстройства, мигрень, начальные стадии гипертонической болезни и атеросклероза, энцефалопатии разного происхождений также в начальных своих стадиях, Сюда же можно отнести больных с отделенными последствиями черепно-мозговой травмы и нейроинфекций (исключая острую травму и свежие нейроинфекции).
вот оказывается, что метод ЭЭГ дает одну из немногих возможностей объективизировать фактическое положение вещей. Если менее нарушена, чем это можно было бы ожидать на основании клинического обследования больного, значит нарушения мозга,
62
органическое ядро заболевания хорошо компенсировано. Если же ЭЭГ нарушена больше, чем это можно было бы ожидать, то значит у пациента один из вариантов болезней регуляции. Показания ЭЭГ служат в этом случае для выбора тактики лечебных мероприятий.
Итак, в тех случаях, которые не относятся к эпилепсии или грубым (макроочаговым) поражением мозга, оценку показаний ЭЭГ следует производить, признавая, во-первых, существование особой формы патологии – болезней регуляций, и во -вторых , постулируя, что большинство регистрируемых на ЭЭГ сдвигов происходит за счет дистантных влияний на кору со стороны многозвеньевой системы лимбико-ретикулярного комплекса.
Важное значение для суждения о патологии мозга имеют реактивные пробы. Умеренно выраженные реакции на открывание и закрывание глаз и световые раздражения свидетельствуют в пользу достаточной сохранности всех структур мозга. Усиление в ответ на реактивные пробы выраженности патологических компонентов ЭЭГ (медленных тота- и дельта -волн, острых волн) происходит обычно в областях коры мозга, окружающих очаги ее поражения. Резко усиленные реакции навязывания ритма световых раздражений указывают на повышенную активность всех структур таламуса (его неспецифических, специфических и ассоциативных ядер ). Ослабление вплоть до полного исчезновения реакций ЭЭГ на пробы с открыванием –закрывание глаз и на действие световых раздражений указывает на функциональную недостаточность и даже на частичное или полное разрушение связей коры с нижележащими отделами мозга.
§ 3.
63
а свете сказанного в §1 и §2, я думаю, что наибольшей информативности метода электроэнцефалографии можно было бы достигнуть, если бы удалось построить модели отдельных электроэнцефалографических синдромов. Попытки такого рода имеются в литературе.
Так П. А. Жоров [16] пишет, не называя их синдромами, о четырех взаимоисключающих вариантах биоэлектрической активности мозга.
Первый вариант «корковый». Он обусловлен высокой активностью коры и относительно низкой активностью подкорки. На ЭЭГ при этом альфа-ритм высокого индекса и высокой амплитуды, но относительно малой частоты. Кожно-гальваническая реакция в этом варианте имеет небольшую интенсивность и быстро угасает.
Второй вариант «корково-подкорковый». Он связан с высокой активностью, как коры, так и подкорки. На ЭЭГ альфа-активность относительно малой частоты и обильная бета-активность, усиленная по индексу и амплитуде. Кожно-гальваническая реакция высокой интенсивности, медленно угасающая.
Третий вариант «подкорково-корковый». Он отражает низкую активность, как коры, так и подкорки. На ЭЭГ альфа-активность малой частоты. Кожно-гальваническая реакция низкой интенсивности, медленно угасающая.
Четвертый вариант «подкорковый». Он характеризуется высокой активностью подкорки и низкой активностью коры. На ЭЭГ альфа-активность большой частоты и усиление интенсивности бета-колебаний.
Работа эта в известном смысле примитивна. Сложные мозговые структуры фигурируют в ней лишь в виде общих
64
понятий – кора и подкорка. В биоэлектрической активности мозга учитываются лишь два компонента – альфа- и бета- колебания , и их описание дается весьма упрощенно. Но сама идея системного подхода при оценке паттернов ЭЭГ заслуживает признания. Рассуждения автора можно было бы модернизировать, уточнив анатомические характеристики мозга и расширив число описываемых компонентов ЭЭГ. Безусловным прогрессом
и системном подходе к анализу ЭЭГ является монография В. В. Трошина [48], излагающая результаты изучения черепно-мозговой травмы. Исследования Трошина выполнены на высоком профессиональном уровне. Подробно и компетентно описаны все изменения биопотенциалов при разных стадиях легкой и тяжелой травмы. Различаются 11 вариантов сдвигов на ЭЭГ, которые называет типами ЭЭГ. Показывается встречаемость каждого из типов при разных случаях травмы. Особый интерес вызывает заключительная часть
монографии, где автор вводит понятие электроэнцеграфического синдрома. Описывается 4 синдрома:
Синдром активации ЭЭГ. Преобладает бета-
активность высокой частоты с амплитудой от 5 до 30 мкв. На этом фоне в задних отделах мозга-нерегулярная альфа-активность с амплитудой до 50 мкв, а в передних отделах – нерегулярная медленная активность с амплитудой до 30 мкв. Вспышки на ЭЭГ отсутствуют,
Синдром начальной синхронизации ЭЭГ. Альфа-
ритм нерегулярный по частоте без амплитудных модуляций. Много тета- и дельта-волн (одиночных или группами) с амплитудой 50-70 мкв. В 25% случаев встречаются «вспышки»: внезапно появляющиеся и
65
исчезающие группы из самых разных комбинаций полиморфных бета-, альфа-, тета- и дельта-волн в 1,5-3 раза превышающих по амплитуда фоновую активность. Могут иметь место очаговые нарушения на ЭЭГ (24% случаев). Реакции на афферентные раздражения обычно сохранны.
Синдром выраженной синхронизации ЭЭГ.
Значительная дезорганизация и замедление альфа-ритма, на который накладывается бета-активность. Обильная медленная активность с амплитудой до 110 мкв. Наблюдается большое количество вспышек (37% случаен)
В очаговые нарушения (7% случаев). Реакции на афферентные раздражения часто отсутствуют или ослаблены.
Синдром максимальной синхронизации ЭЭГ. Доминируют тета-и дельта-ритмы с амплитудой более 110 мкв без особых зональных различий. Вспышки встречаются очень часто (91% случаев), а очаговые нарушения реже (35% случаев).
Различия между синдромами автор трактует как отражение влияний на кору двух отделов неспецифической системы мозга – стволового и лимбического. Эти отделы находятся в реципрокных отношениях. При возбуждении стволовых структур активность лимбического отдела угнетается, а при возбуждении лимбического отдела влияния со стороны среднего мозга блокируются на уровне гипоталамуса. В это время из лимбической системы в кору посылаются синхронизирующие влияния.
Высоко оценивая монографию Трошина в целом, нельзя не высказать некоторые критические замечания. Прежде всего характеристика отделов неспецифической системы, как морфологическая, так и функциональная
66
слишком схематична. Как это было показано нами выше, современные представления по этому вопросу раскрывают гораздо большую сложность как строения, так и функций разных отделов лимбико- ретикулярного комплекса. Схематичность представлений автора находит отражение и
\endash формировании выделенных им синдромов. Только один синдром автор считает отражением активирующих влияний, а три других как проявления синхронизирующих влияний. По сути дела эти три синдрома различаются лишь по степени изменений ЭЭГ. Но ведь понятие «синдром» предполагает качественные, а не количественные различия. Наоборот, выделяемые автором критерии для кодирования ЭЭГ слишком усложнены, а построенная классификация ЭЭГ недостаточно логична. В ней нет стройности, нет выделения какого-либо градиента нарастания или убывания сдвигов на ЭЭГ. Я много лет занимаюсь типологическими исследованиями ЭЭГ и, как мне кажется, нашла более удачный вариант классификации, который будет изложен позднее.
Наиболее строгий синдромологический подход к трактовке феноменов ЭЭГ мы находим в интереснейшей монографии Н. Н. Траугатт с коллективом соавторов [47]. Монография посвящена подробному изучению механизмов действия пяти фармакологических агентов (аминазин, инсулин, амитал натрия, тофранил, бемегрид). Исследования проводились в психиатрической клинике в процессе лечения больных. Наряду о фиксацией клинических данных, изучались: ряд инструментальных показателей состояния сердечно-сосудистой системы; показатели состояния высшей нервной деятельности (условные и безусловные рефлексы); динамика изменений биоэлектрической активности мозга; электрическая
67
активность мышц и кожно-гальваническая реакция. Анализ полученных данных авторы производили в свете представления о том, что особенности целостной реакции организма на фармакологические воздействия зависят от того, какие неспецифические структуры лимбико-ретикулярного комплекса раздражаются или угнетаются
применяемыми фармакологическими агентами. Учитывалось состояние следующих шести структур: ретикулярной формации ствола мозга; неспецифических ядер зрительного бугра; ассоциативных ядер зрительного бугра; заднего гипоталамуса; переднего гипоталамуса; хвостатого ядра. В таблице, составленной нами по данным, приведенным в монографии, показывается, в каком состоянии, в соответствии с известными фармакологическими свойствами каждого агента и в зависимости от стадии действия агента, должна находиться каждая из мозговых структур. Целостную совокупность всех изменений в каждом из изученных случаев авторы
называют КЛИНИКО-ФИЗИОЛОГИЧЕСКИМ СИНДРОМОМ (табл. №2).
СВОДНАЯ ТАБЛИЦА , СОСТАВЛЕННАЯ ПО ДАННЫМ ТРАУГОТТ с соавт. Таблица №2
| Аминазин | Инсулин | Тофранил | Барбамил | Бемегрид | ||||
| Первая | Вторая | Третья | Вторая | Третья | ||||
| фаза | фаза | фаза | фаза | фаза | ||||
| Ретикулярная | - | + | - | - | + | + | - | - |
| формация ствола | ||||||||
| Неспецифические | - | - | + | - | + | - | + | + |
| ядра таламуса | ||||||||
| Ассоциативные | 0 | 0 | 0 | - | + | + | - | + |
| ядра таламуса | ||||||||
| Задний | - | + | - | - | + | + | - | - |
| гипоталамус | ||||||||
| Передний | + | 0 | 0 | + | - | 0 | 0 | 0 |
| гипоталамус | ||||||||
| Хвостатое ядро | - | 0 | 0 | 0 | 0 | 0 | + | 0 |
В состояние угнетения
у состаяние раздражения
4. спокойное «рабочее» состояние
68
§ 4.
Опираясь на факты, представленные в фундаментальном исследовании Н. Н. Трауготт с соавторами, мы сделали попытку выделения различных электроэнцефалографических синдромов. Эти синдромы имеют сходство с клинико-физиологическими синдромами, описанными авторами монографии, но не повторяют их. Названия наших синдромов отражают феноменологию ЭЭГ, а не механизмы их формирования.
2. Синдром гиперсинхронизации альфа-активности – условное обозначение «αα».
Модель – вторая стадия действия инсулина. На ЭЭГ усиление альфа-активности по индексу и амплитуде, стирание (или сглаживание) зональных различий (распространение альфа-активности на передние отделы мозга). Иногда на этом фоне возможны короткие вспышки дельта-волн высокой амплитуды, главным образом, в лобных отделах мозга. Вспышки усиливаются при гипервентиляции. Все электрографические реакции на афферентные раздражения ослаблены.
Механизм формирования синдрома. Угнетение активности ретикулярной формации ствола мозга и заднего гипоталамуса. Усиление деятельности неспецифического таламуса за счет его высвобождения от тормозных влияний со стороны ретикулярной формации ствола. Спокойное (рабочее) состояние переднего гипоталамуса, ассоциативных ядер таламуса и хвостатого ядра (Рисунок №2).
Клинически: вялость, сонливость, переходящая в оглушение! В вегетативно-сосудистой системе усиление парасимпатических влияний.
3. Синдром десинхронизации альфа-активности -
69
условное обозначение β2.
Модель – действие адреналина, третья стадия действия тофранила и (в известной степени) первая стадия действия инсулина.
На ЭГГ по всем областям мозга доминирует бета-активность высокой частоты, малой амплитуды. При афферентных раздражениях усиление феномена десинхронизации. Реакция усвоения (навязывания) ритма световых раздражений ослаблена или отсутствует.
Механизм формирования синдрома. Угнетение активности неспецифических ядер зрительного бугра. Усиление деятельности ретикулярной формации ствола и заднего гипоталамуса (иногда усиливается также деятельность ассоциативных ядер таламуса). Спокойное, (рабочее) состояние переднего гипоталамуса и хвостатого ядра (Рисунок №3).
Клинически: раздражительность, тревога, усиление симпатических эффектов (повышение артериального давления, снижение порога вегетативных эффектов).
70
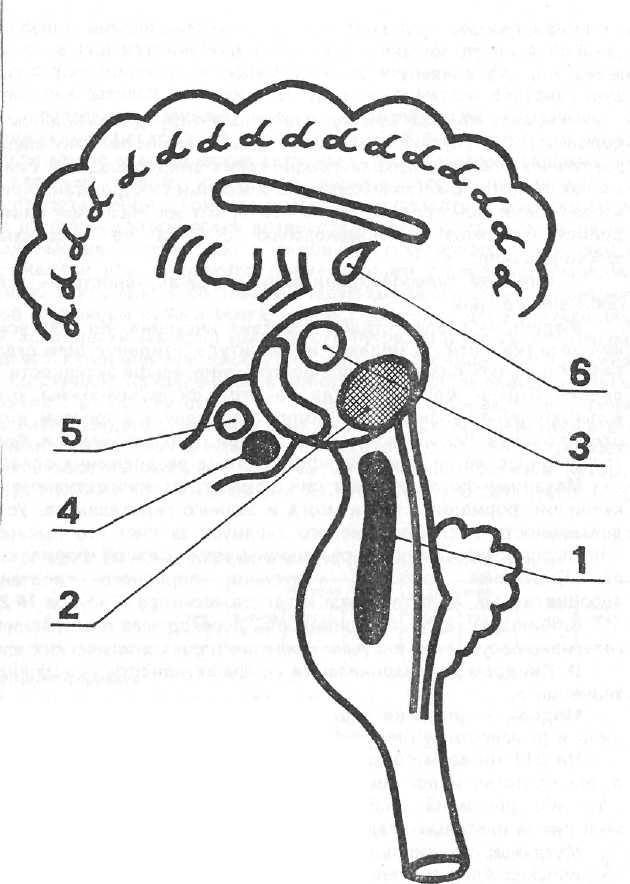
Рисунок №2. Модель синдрома 1
1 Ретикулярная формация ствола мозга; 2 Неспецифические ядра таламуса; 3. Ассоциативные ядра таламуса; 4. Задний гипоталамус; 5.
Передний гипоталамус; 6. Хвостатое ядро
 состояние раздражения
состояние раздражения
состояние угнетения
71
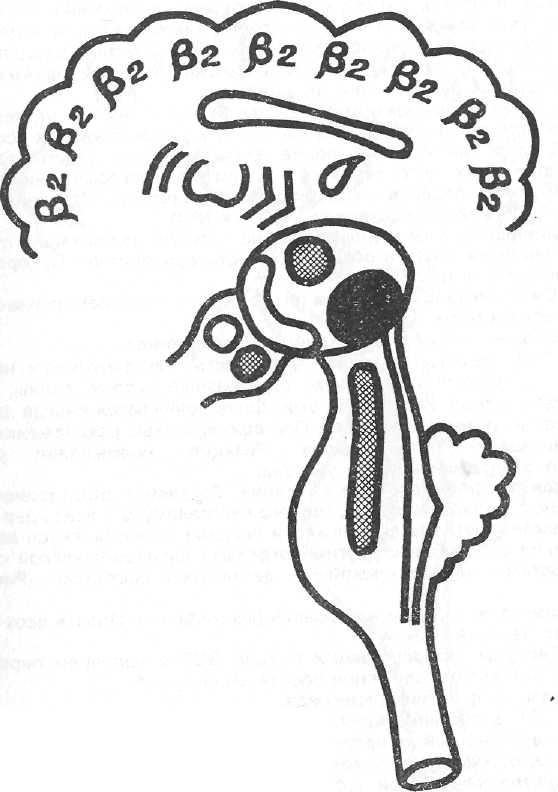
Рисунок №3. Модель синдрома 2
Обозначения как на рис. 2
72
2. Синдром гиперсимхронизации бета-активности – условное обозначение β1
Модель – действие амитала натрия.
На ЭЭГ по всем областям мозга доминирует бета-активность низкой частоты (14-24 пер/сек), нередко имеющая высокую амплитуду (до 40 мкв и более), иногда группирующаяся в веретена. Возможна примесь групп альфа-волн (8-12 пер/сек), также иногда в виде веретен, преимущественно в лобных отделах. Афферентные раздражения могут усиливать амплитуду биопотенциалов. Реакция навязывания ритма световых раздражений не выражена.
Механизм формирования синдрома. Резкое угнетение деятельности ретикулярной формации ствола, заднего гипоталамуса и ассоциативных ядер таламуса. Усиление активности хвостатого ядра и неспецифических ядер таламуса вследствие высвобождения их от влияний других отделов неспецифической системы. Передний гипоталамус
и спокойном состоянии (Рисунок № 4).
Клинически: успокоенность, снятие тревоги, напряжения, страха , иногда эйфория. Снятие обездвиженности при ступоре. Судорожная готовность мозга низкая.
и Синдром дезорганизации ритмов ЭЭГ с наличием вспышек. Условное обозначение β2wα.
Модель – вторая стадия действия тофранила.
На ЭЭГ дезорганизованная активность. Регистрируется нерегулярный по частоте альфа-ритм, однофазные острые волны, бета-активность разной частоты. На этом фоне появляются иногда вспышки из острых и медленных волн. При афферентных раздражениях десинхронизация биопотенциалов. Реакция
73
навязывания ритма световых раздражений резко усилена. Механизм формирования синдрома. Состояние
раздражения ретикулярной формации ствола, заднего гипоталамуса и всех ядер таламуса . Резкое угнетение деятельности переднего гипоталамуса за счет разобщения его связей с другими отделами неспецифической системы. Хвостатое ядро в спокойном, деятельном состоянии. (Рисунок №5).
Клинически: хорошее настроение без эйфории.
Пороги всех вегетативных реакций низкие.
с Синдром дезорганизации ритмов ЭЭГ с наличием пароксизмальной активности. Условное обозначение
β2wα↑.
Модель – действие бемегрида.
На ЭЭГ дезорганизованная активность. Регистрируется нерегулярный, замедленный по частоте альфа-ритм, иногда увеличенный по амплитуде; однофазные острые волны, мелкие пики; бета-активность преимущественно высокой частоты. В целом паттерн имеет сходство с таковым в синдроме 4. Отличие в том, что на этом фоне выражены не вспышки, а высокоамплитудные пароксизмальные разряды тета- и дельта- волн. Кроме того, десинхронизация биопотенциалов при действии афферентных раздражений ослаблена. Однако реакция навязывания
74
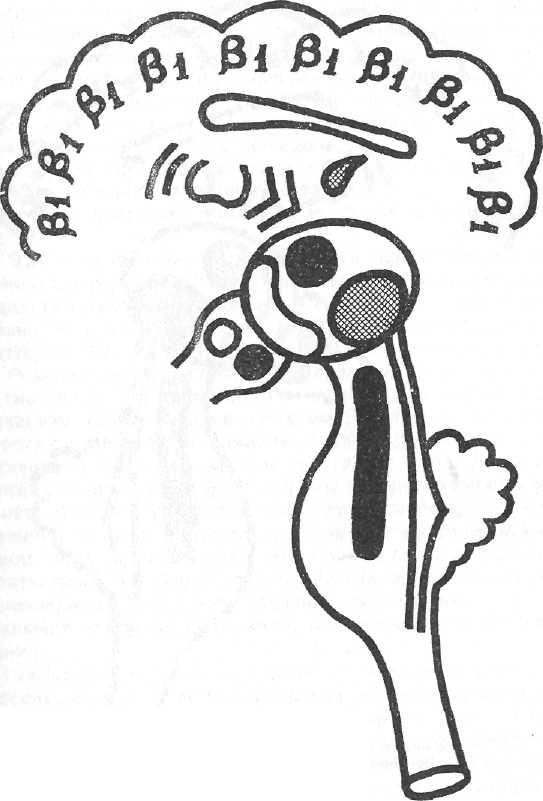
Рисунок № 4. Модель синдрома 3.
Обозначения как на рис. 2
75
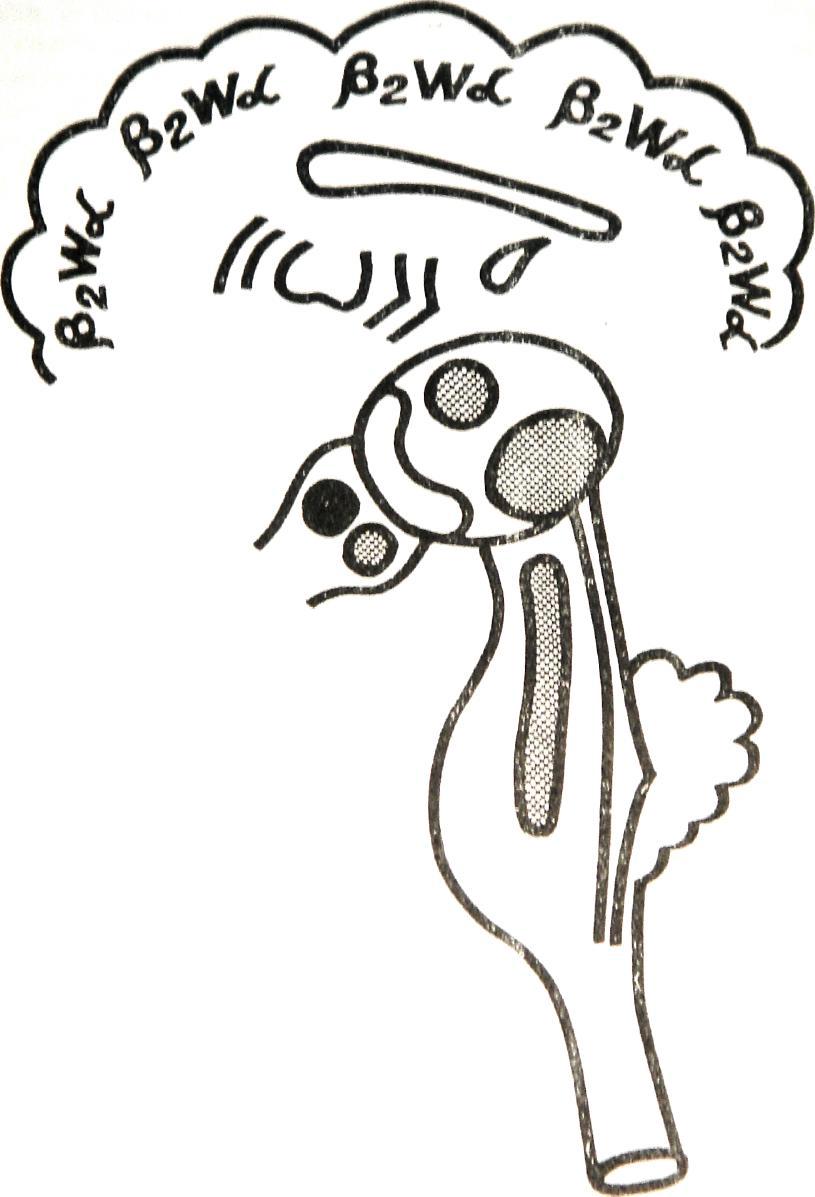
Рисунок №5, Модель синдрома 4.
Обозначения как на рис. 2
76
ритма световых раздражений, как и в предыдущем синдроме, резко усилена.
Механизм формирования синдрома. При состоянии раздражения как специфических, так и ассоциативных ядер таламуса, угнетение деятельности ретикулярной формации ствола мозга и заднего гипоталамуса. Передний гипоталамус и хвостатое ядро в спокойном состоянии. (Сходные механизмы формирования паттерна ЭЭГ действуют и при функциональной пробе с гипервентиляцией (Рисунок №6).
Клинически: хорошее настроение, снятие тревоги, речевой и двигательной обездвиженности. Пороги вегетативных реакций высокие. Судорожная готовность мозга повышена.
[72] Синдром перестройки корковой ритмики с замедлением биопотенциалов. Условное обозначение θΔ.
Модель – действие аминазина и третья стадия действия инсулина.
На ЭЭГ – ослабление бета-активности, ослабление вплоть до исчезновения альфа-активности. Появление и все большее усиление тета- и дельта-активности. Иногда вспышки еще более выраженных медленных волн высокой амплитуды в лобных отделах мозга. (Сходные паттерны ЭЭГ могут наблюдаться при гипогликемии и гипоксии мозга). Реакция на афферентные раздражения и реакция навязывания ритма световых раздражений ослаблены или отсутствуют.
Механизм формирования синдрома. Глубокое подавление деятельности ретикулярной формации ствола, заднего гипоталамуса, неспецифических ядер таламуса и хвостатого ядра. Доминирование восходящих влияний на кору со стороны переднего гипоталамуса, деятельность
77
которого усилена за счет его высвобождения от тормозных влияний со стороны других отделов неспецифической системы.
Клинически: эмоциональное безразличие, глубокая успокоенность без утраты сознания. Своеобразный, неглубокий, легко прерываемый сон. Снижение артериального давления, гипотермия.
Заключая описание синдромов, необходимо сделать некоторые замечания:
Поскольку синдромы выделены на основании фармакологических исследований в психиатрической клинике, желательно) подтверждение значимости описанных данных при проведении исследований среди больных другого профиля, при проведении терапии иной
природы (мамритор, рефлексотерапии или иглоукалывания).
Нами пока выделены только те синдромы, обоснование которым мы нашли в исследованиях Трауготт
В соавторами. Безусловно можно и нужно выделить и другие электроэнцефалографические синдромы, например, ЭЭГ с нормально организованным альфа-ритмом; «плоская» ЭЭГ; ЭЭГ с локальными очаговыми нарушениями и т. д.
78
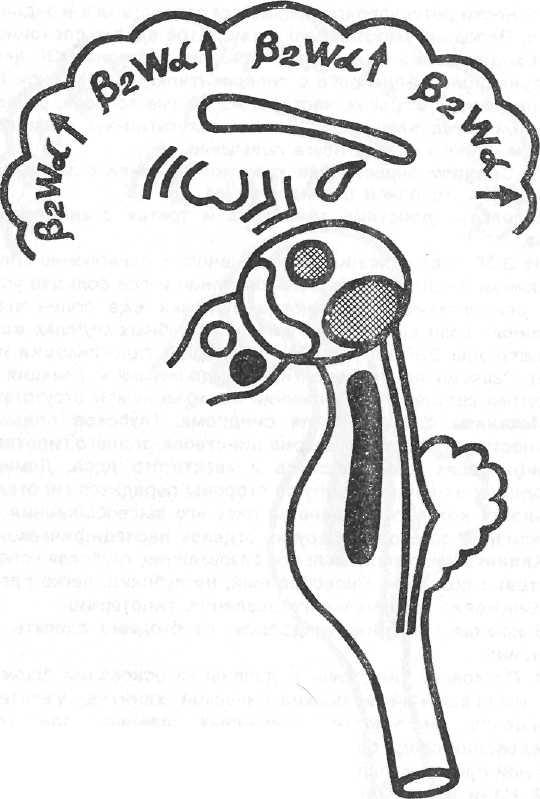
Рисунок 5. Модель синдрома 5. Обозначения как на рис.2.
79
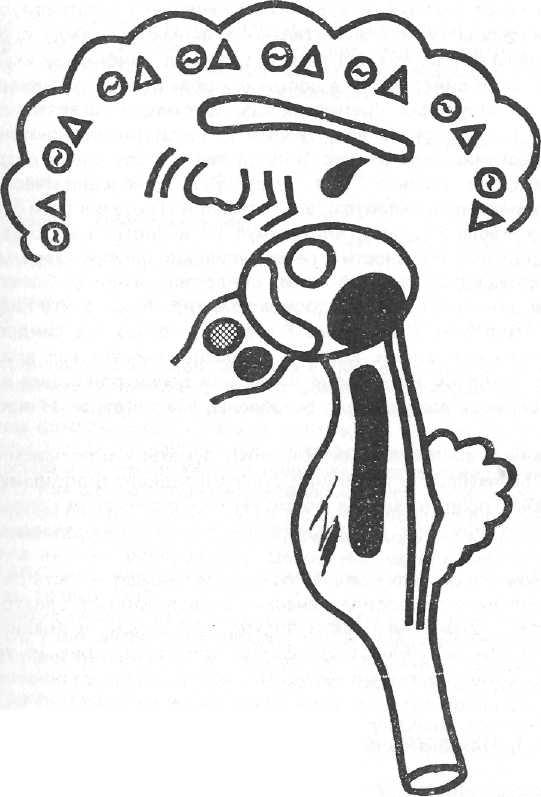
Рисунок № 7. Модель синдрома 6. Обозначения как на рис. 2
80
О Выделенные синдромы выявляют некоторые интересные закономерности.
а) Синдром 3 подчеркивает связь появления на ЭЭГ бета-активности низкой частоты с повышением активности хвостатого ядра.
б) Синдром 6 иллюстрирует зависимость замедления ритмов биопотенциалов от влияний, исходящих из переднего гипоталамуса.
в) Синдромы IV и V значительно различаются между собой по функциональному состоянию разных разделов лимбико-ретикулярного комплекса. В синдроме IV в состоянии повышенной активности находятся 4 структуры (ретикулярная формация ствола, задний гипоталамус и две группы ядер таламуса), а в состоянии угнетения находится передний гипоталамус. В синдроме V в состоянии раздражения находятся только две структуры (неспецифические и ассоциативные ядра таламуса), а в состоянии угнетения ретикулярная формация ствола и задний гипоталамус. Но несмотря на это, характеристика фоновой активности и резкое усиление реакции навязывания ритма световых раздражений в этих синдромах сходные. Электрографическим различием двух синдромов является лишь то, что пароксизмальная активность в синдроме 5 выражена резко, а в синдроме 4 представлена вспышками, которые не расцениваются как эпилептиформные. При этом в синдроме 5 реакции десинхронизации в ответ на афферентные раздражения ослаблены, а в синдроме 4 наоборот усилены.
Указанные сопоставления позволяют думать, что повышенная активность ретикулярной формации ствола и заднего гипоталамуса может служить препятствием к развитию эпилептических проявлений, которые, по-
81
видимому, провоцируются со стороны таламических структур мозга.
с целом все изложенное в §4 показывает
желательностьрасширенияиуглубления
синдромологических исследований в электроэнцефалографии. Думаю, что такой подход к описанию и анализу ЭЭГ способен вывести клиническую электроэнцефалографию из того тупика, в котором она сейчас находится.
Дата: 2019-03-05, просмотров: 406.