1. Поступательное - обусловлено тем, что на пути следования возбуждения встречается тормозной нейрон.
2.Возвратное - осуществляется клетками Реншоу для подаволения избыточной активности мотонейронов. Аксоны мотонейронов часто дают коллатерали (ответвления), оканчивающиеся на клетках Реншоу, аксоны которых, в свою очередь, оканчиваются на теле или дендритах тех же мотонейронов, образуя тормозные синапсы. Возбуждение, возникающее в мотонейроне, распространяется по прямому пути к скелетной мускулатуре, а также по коллатералям к тормозящему нейрону, который посылает импульсы к мотонейронам и тормозит их. Чем сильнее возбуждение мотонейрона, тем сильнее возбуждаются клетки Реншоу и тем более интенсивно они оказывают тормозящее действие, подавляя чрезмерную активность мотонейронов.
3. Латеральное - вставочные клетки формируют тормозные синапсы на соседних нейронах, блокируя боковые пути распространения возбуждения. В таких случаях возбуждение направляется по строго определенному пути. Этот вид торможения обеспечивает направленную иррадиацию возбуждения в ЦНС.
4. Реципрокное - например, торможение центров мышц-антагонистов. Суть данного вида реципрокного торможения состоит в том, что возбуждение проприорецепторов мышц-сгибателей одновременно активирует мотонейроны данных мышц и вставочные тормозные нейроны. Возбуждение вставочных нейронов приводит к постсинаптическому торможению мотонейронов мышц-разгибателей.
Динамика и взаимодействие процессов возбуждения и торможения
Движение нервных процессов:
1. Иррадиация - распространение из какого-либо очага или нервного центра на окружающие нервные клетки процесса возбуждения или торможения. В случае иррадиации возбуждения в ответной реакции принимает участие не одиночный нервный центр, а вся нервная система и организм в целом. Иррадиация возбуждения и торможения возникает лишь в ответ на сильные и длительно действующие стимулы. Нервный процесс средней силы не иррадирует или распространяется в пределах нескольких нервных центров.
2.Концентрация - стягивание распространившегося возбуждения или торможения к группе определенных клеток одного нервного центра, т. е. к исходному пункту, откуда оно началось, усиливая его положительную или отрицательную функцию. Примером концентрации возбуждения может быть процесс специализации условного рефлекса, когда возбуждение сосредоточивается в одном нервном центре, обеспечивая точность ответного действия на условный раздражитель.
3.Индукция - возникновение вокруг очага одного из процессов распространения другого процесса.
Положительная индукция - возникновение возбуждения в нервных центрах коры головного мозга вокруг очага торможения.
Отрицательная индукция - возникновение торможения в нервных центрах коры головного мозга вокруг очага возбуждения
Нервные центры
Общие представления о нервных центрах в ЦНС
Нервные центры
-в узком смысле: это совокупность нейронов, принимающих участие в осуществлении конкретного рефлекса (мигания, глотания, кашля и т. д.);
- в широком смысле: функциональное объединение нейронов, расположенных на различных уровнях ЦНС при формировании сложного адаптивного поведения.
Свойства нервных центров
· Одностороннее проведение возбуждения. Возможно благодаря существованию межнейронных синапсов. Синапсы передают возбуждение только в одном направлении, а именно от пре- к постсинаптической мембране, обеспечивая передачу информации строго с афферентных систем на эфферентное.
· Центральная задержка. Общее время проведения возбуждения определяется силой возбуждения и протяженностью рефлекторной дуги, а также свойствами ее нейронов и синапсов. Из всех элементов рефлекторной дуги, участвующих в проведении импульсов, медленнее всего возбуждение проводится через нервные центры. Это происходит вследствие наличия в нервных центрах большого числа синапсов, обуславливающих синаптическую или центральную задержку. Центральная задержка связана с большим числом вставочных нейронов в нервных центрах. Большое число синапсов удлиняет время проведения возбуждения в силу временных затрат на выделение медиатора, взаимодействие медиатора с рецептором, формирование потенциала действия.
· Трансформация ритма. Частота импульсов на входе и выходе из нервных центров может быть неодинаковой: нервные центры могут менять частоту импульсов (трансформировать частоту). (1) Снижение частоты импульсов - увеличение рефрактерного периода в нервных центрах и соответствующее в связи с этим уменьшение лабильности. (2) Обратная трансформация, то есть увеличение частоты импульсации. Смена редких импульсов на более частые связана во-первых, с задержкой следовой электроотрицательности, что ведет к появлению очередного потенциала действия; во-вторых, задержка во времени постсинаптической деполяризации, что предоставляет возможность сформироваться нескольким дополнительным потенциалам действия.
· Последействие - сохранение возбуждения после того как раздражитель уже перестал действовать на центр. Выделяют кратковременное и долговременное последействия. Описано два основных механизма последействия. Во-первых, это задержка постсинаптической деполяризации, во-вторых - циркуляция импульсов по замкнутым нервным цепям, т. е. их реверберация. Благодаря существованию этих замкнутых кольцевых нервных связей возбуждение долго сохраняется в нервных центрах.
· Суммация возбуждения. Впервые была описана И. М. Сеченовым в 1863 году. Может быть последовательной или одновременной (пространственной). Последовательная суммация происходит в случаях, когда с одного рецепторного поля к нервному центру через короткий интервал времени поступают два возбуждения, причем суммированное возбуждение больше по величине, чем арифметическая сумма отдельно взятых двух возбуждений (отмечается эффект потенциирования). Суммация возбуждения наблюдается при действии раздражителей различной силы, особенно хорошо же видна при действии двух подпороговых раздражителей. Если два подпороговых раздражения подавать в отдельности, то возбуждение нервного центра не наблюдается, а когда они следуют друг за другом - происходит. Механизм последовательной суммации представляет собой накопление пороговой концентрации нейромедиатора на одних и тех же синапсах (медиатор, выделившийся вследствие действия первого подпорогового стимула, не успевает разрушиться). Одновременная суммация(пространственная) - суммация одновременных импульсов, поступивших с разных участков одного рецепторного поля, при этом общий эффект также превышает простую арифметическую суммы отдельно взятых импульсов.
· Посттетаническая потенциация - повышение возбудимости центра, сопровождающееся значительным увеличением амплитуды синаптических потенциалов, после поступления частых импульсов (более 400-500 имп/с). После очень продолжительной стимуляции уровень посттетанической потенциации может возрасти во много раз, а ее продолжительность может составлять от нескольких минут до нескольких часов. Посттетаническая потенциация относится к процессам облегчения в ЦНС, задействована в процессах памяти и обучения.
· Конвергенция – импульсы, приходящие в ЦНС по различным эфферентным волокнам, могут конвергировать к меньшему количеству промежуточных и афферентных нейронов. Это явление легло в основу принципа конвергенции, установленного Шеррингтоном. Конфергенция связана с процессами фильтрации информации.
· Дивергенция – возбуждение в ЦНС может распространяться от одного нейрона на несколько, в этом случае присходит усиление. Свойство является обратным конвергенции..
· Тонус нервных центров - постоянное возбуждение нервных центров в состоянии покоя, без внешней стимуляции. Полагают, что тонус нервных центров поддерживается нервными импульсами, непрерывно поступающими от рецепторов (особенно проприорецепторов) и различными гуморальными влияниями (гормоны, СО2 и др.).
· Утомление нервных центров - нервные центры легко утомляемы ( в то время как для нервных путей утомление не характерно), утомление нервного центра проявляется постепенным снижением, а затем и полным прекращением рефлекторного ответа.
· Высокая чувствительность к недостатку кислорода и действию ядов.
· Центральное возбуждение - центральное возбуждение возникает при стимуляции нервных центров и распространяется (иррадиирует) на другие центры. Иррадиация может носить избирательный характер, когда возбуждение распространяется в определенном направлении, и диффузный - возбуждение распространяется равномерно, на все участки. Состояние диффузной иррадиации возбуждения нервных центров свойственно новорожденным и детям первых месяцев жизни, т. к. нейроны и нервные волокна еще недостаточно сформировались, например, полностью не миелинизировались.
· Центральное торможение было открыто в 1862 г. И.М. Сеченовым. В нервных центрах при действии раздражителей наблюдается центральное торможение, которое проявляется подавлением функциональной активности какого-либо органа или системы. Доказательством наличия в ЦНС торможения явились классические опыты И. М. Сеченова: у лягушки с перерезаным головным мозгом на уровне зрительных бугров на место среза помещался кристаллик поваренной соли. При погружении задней лапки лягушки в слабый раствор серной кислоты время рефлекса удлинялось, а после удаления раздражителя оно восстанавливалось. И.М. Сеченов заключил, что на уровне ствола мозга находятся тормозные структуры, угнетающие спинномозговые рефлексы. Позже было показано, что данные тормозные структуры принадлежат к ретикулярной формации.
· Пластичность нервных центров — способность перестраивать функциональные свойства для более эффективной регуляции функций, осуществления новых, ранее несвойственных этому центру рефлексов или восстановления функций после повреждения части нейронов центра. Пластичность обеспечивает изменение эффективности и направленности связей между нервными клетками, является рабочим механизмом обучения. В основе пластичности лежат функциональные особенности синапсов и мембран нейронов («тренировка» синапсов, посттетаническая потенциация, периодичность функционирования синапсов и нейронов, пространственная и временная суммация постсинаптических потенциалов), а также наличие многочисленных дублирующих систем нейронов и нервных волокон.
· Целостные формы поведения
· Целостные формы поведения
· Любое поведение может являться адаптивным, только если оно согласовано на уровне всего организма и адекватно по отношениям как к собственным потребностям, так и к условиям внешней среды.
· Американцам пришла в голову простая идея: разъединить правое и левое полушария головного мозга, рассечь нервные связи между ними, чтобы предотвратить систематическое распространение эпилептических разрядов на весь мозг. Полушария соединены миллионами нервных волокон, которые передают информацию из одного полушария в другое и образуют так называемое мозолистое тело — белесоватую плотную массу, создающую как бы мост между двумя полушариями. Такая операция была произведена на нескольких больных, она действительно облегчила их страдания и одновременно привела к крупнейшему открытию, удостоенному в 1980 г. Нобелевской премии. Ее получил Р. Сперри.
· Что же произошло после рассечения мозолистого тела с поведением и психикой человека? На первый взгляд, ничего особенного, и это уже было достаточно удивительно. Связи между двумя половинами мозга были разрушены, а человек ел, совершал повседневные поступки, ходил и беседовал с другими людьми без серьезных видимых отклонений в поведении. Правда, настораживали несколько наблюдений, сделанных вскоре после операции: один пациент пожаловался, что он странно ведет себя с женой и не в состоянии контролировать свое поведение: в то время как его правая рука обнимает жену, его левая рука ее отталкивает. Другой пациент обратил внимание на странное поведение своей левой руки перед посещением врача: в то время как с помощью правой руки он одевался и приводил себя в порядок, левая рука пыталась расстегнуть и снять одежду. Возникала ситуация, описанная в метафоре, когда левая рука не знает, что делает правая. Дело, однако, было не в руках: это одна половина мозга не знала, что делает другая половина.
· Роттенберг В.С. Сновидения, гипноз и деятельность мозга.
·
· Для формирования целостного поведения необходимо следующее.
· 1). Адекватно определять собственные актуальные потребности и оценивать текущие внешние условия.
· 2).Обеспечивать уровень энергетического обеспечения ЦНС и организма в целом, ориентируясь на стоящие задачи.
· 3). Формировать и осуществлять программу действия, обеспечивая координацию работы всех частей организма.
· 4). Подстраивать поведение под меняющуюся среду, используя механизм обратной связи; отслеживать результаты поведения и вносить коррекцию в программу действия.
· Одна из теорий, которая объясняет механизмы формирования целостного поведения - теория функциональных систем К. Анохина.
· 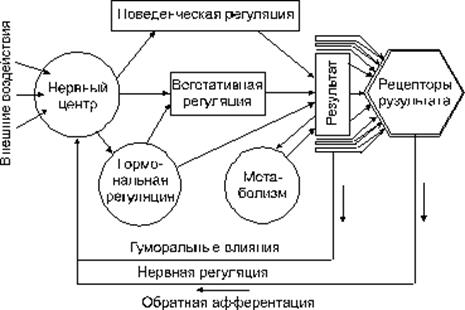
· Рис.1 Регуляторные механизмы в функциональной системе
· 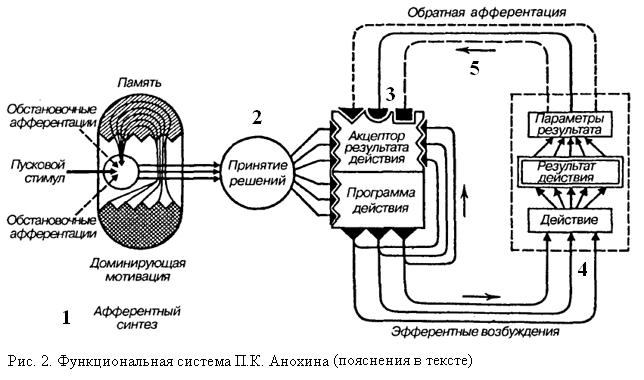
· Функциональная система - единица интегративной деятельности организма, представляющая собой динамическую морфофизиологическую организацию центральных и периферических образований, избирательно объединенных для достижения полезного для организма приспособительного результата.
· Функциональный системы обладают способностью экстренной самоорганизации за счет быстрой мобилизуемости взаимодействующих компонентов. Возникают для удовлетворения возникшей потребности, при формировании образа конечного результата его удовлетворения, то есть построены по принципу ориентации на цель.
· Для того, чтобы достигнуть результата, функциональная система осуществляет ряд процессов, описанных ниже.
· 1). Афферентный синтез всей поступающей в центральную нервную систему. информации.
· 2) Принятие решения с одновременным формированием аппарата прогнозирования результата в виде афферентной модели — акцептора результатов действия.
· 3) Реализация принятого решения в действии.
· 4) Сличение афферентной модели акцептора результатов действия и параметров результатов выполненного действия, полученных организмом при помощи обратной афферентации.
·
· Формирование функциональной системы начинается с афферентного синтеза, в процессе которого происходит взаимодействие мотивационного возбуждения, обстановочной афферентации и извлеченных из памяти следов образов прошлого опыта. После обработки всей совокупности данной информации принимается решение о конкретных действиях и происходит переход от обработки информации к формированию последовательной программы действия.
· Под влиянием пускового стимула скрытая предпусковая интеграция в виде команды, представленной комплексом эфферентных возбуждений, направляется к периферическим органам и реализуется в соответствующем действии. Параллельно с формированием программы действия формируется акциптор результатов действия, направленный на считывание (путем обратной афферентации) информации о результатах.
· Данная информация сопоставляется с афферентной моделью результата, хранящейся в акцепторе действия. Совпадение заготовленного образа и того, который возникает при считывании результатов, является сигналом успеха действия, и организм переходит к следующему действию (формируется новая функциональная система). Несовпадение модели акцептора действия с обратной афферентацией, т. е. рассогласование, вызывает ориентировочно-исследовательскую реакцию, новый афферентный синтез с подбором информации, необходимой для принятия решения, соответствующего изменившейся обстановке.
· Помимо осуществления единичного действия, целостность поведения реализуется и за счет создания сложных последовательных комплексов действий. На уровне врожденного поведения это - инстинктивные формы поведения, предполагающие сложную последовательность действий.
· В частности, Н. Тинберген разработал иерархическую теорию инстинкта, в рамках котрого предполагается, что существует строго координированная и иерерхически организованная система центров мотивации. Удовлетворение потребности, задаваемой однимиз центров, активизирует следующий центр.
· Основой для разработанной Тинбергеном модели поведения послужили следующие факты. Известно, что между различными стереотипными двигательными реакциями существует ряд закономерных отношений. В некоторых ситуациях группы инстинктивных движений появляются совместно — они характеризуют определенное внутреннее состояние животного и проявляют общие флюктуации порога поведенческой реакции. Повышение порога реакции А поднимает порог реакции В (и наоборот), а это свидетельствует о том, что обе они зависят от общего функционального «центра». Наблюдая за сложными поведенческими комплексами действий, можно видеть некоторую регулярность в последовательности проявления тех или иных действий. В качестве примера можно привести агрессивные столкновения рыб за раздел территории. У многих костистых рыб, в том числе и у цихлид, им почти всегда предшествует демонстрация запугивания. Причем у одних видов эти столкновения следуют за очень коротким периодом запугивания; а у других — за весьма разнообразными демонстрациями запугивания, серьезное агрессивное столкновение с ранениями следует лишь в том случае, если силы обоих самцов равны. Наконец, у третьей группы видов настоящие драки уже не наблюдается, и крайне ритуализированная церемония запугивания выполняется до полного истощения одного из соперников, что и решает спор.
· В такого рода ритуализованных столкновениях имеется специфическая последовательность движений: они начинаются с демонстрации боковых поверхностей тела, за которой следует подъем вертикальных плавников. Затем следуют удары хвостом, которые через посредство боковой линии, воспринимающей изменение давления воды, могут, вероятно, сообщить о силе противника. После этого противники встают друг перед другом, вслед за чем начинаются взаимные толчки с открытой пастью, а у других видов — укусы в открытый рот. Они продолжаются до тех пор, пока один из соперников не устанет, окраска его бледнеет и в конце концов он уплывает.
· Такие ритуализованные драки и агрессивные столкновения — прекрасные примеры специфической последовательности стереотипных двигательных реакций: удары хвостом не начнутся до подъема спинного плавника, а толчки отмечаются только после многих ударов хвостом. По интенсивности демонстрации запугивания и ударов хвоста, опытный наблюдатель может определить, кто победит, и начнутся ли толчки «с открытой пастью» вообще, или же один из соперников просто сбежит до начала «серьезной драки».
· Интерпретируя подобные явления, Тинберген выдвинул гипотезу об иерархии центров, управляющих отдельными поведенческими реакциями (рис.2). Согласно Тинбергену, инстинкт представляет собой завершенную иерархическую организацию поведенческих актов, реагирующую на определенный раздражитель четко координированным комплексом действий. Нужно отметить, что понятие «центр» у Тинбергена является главным образом функциональным и не может быть полностью отождествлено с анатомическим понятием нервного центра, хотя Тинберген и ссылается на известные опыты Хесса. В этих опытах при электрораздражении промежуточного мозга кошек наблюдалось развитие некоторых целостных форм поведения (драк, угроз, сна, пищевого, поведения).
· Согласно представлениям Тинбергена, изменение возбудимости центров под влиянием внешних и внутренних воздействий происходит в определенной последовательности. Сначала повышается возбудимость «центра» поисковой фазы поведения, и голодное животное начинает поиск пищи. Когда пища будет найдена, произойдет «разрядка» центра, стоящего на более низком уровне иерархии, и контролирующего осуществления завершающего акта (поедание пищи).
· Схему иерархии центров, управляющих поведением самца колюшки в период размножения, Тинберген представляет следующим образом.
· Высший центр репродуктивного поведения самца активизируется увеличенной длиной дня, гормональными и другими факторами. Импульсы из этого центра снимают блок с центра поискового поведения. Разрядка этого центра выражается в поисках условий для постройки гнезда. Когда такие условия (подходящая территория, температура, необходимый грунт, мелководье, растительность) найдены, происходит разрядка центров следующего уровня иерархии и благодаря этому становится возможной постройка гнезда.
· Если на территорию данного самца проникает соперник, то возбудимость центра агрессивного поведения повышается. Результат этого центра агрессивного поведения — преследования и драки с самцом-соперником. Наконец, при появлении самки повышается возбудимость центра полового поведения, и начинается ухаживание за самкой, представляющее собой комплекс фиксированных действий.
· В дальнейшем вопросы иерархической организации поведения изучал Хайнд (1975). Он показал, что хотя в принципе комплекс фиксированных действий большой синицы можно расположить в иерархическую схему, однако не всегда удается сделать это полностью, так как некоторые из движений характерны для двух и более видов инстинктов. Иногда эти движения являются завершающими актами, а иногда — просто средством создания условий, в которых можно осуществить завершающее действие.
· У молодых животных иерархия поведения часто еще не сформирована. У птенцов, например, сначала появляются на первый взгляд бессмысленные изолированные двигательные акты, и только позже они интегрируются в сложный функциональный комплекс движений, связанных с полетом.
· Расчленение иерархии поведения на элементы часто можно наблюдать во время игры, когда отдельные поведенческие акты, связанные с различными функциями, свободно комбинируются в сочетания не характерных для нормального поведения.
· Существенно, что модель Тинбергена предусматривает возможность взаимодействия между «центрами» различных видов поведения. Дело в том, что случаи, когда животное в каждый данный момент занято каким-то одним видом деятельности, являются скорее исключением, чем правилом. Обычно одни виды активности сменяют другие. Наиболее простой пример такого взаимодействия — подавление одних центров другими. Например, если у самца чайки во время ухаживания за самками усиливается голод, то он может прекратить брачные демонстрации и отправиться на поиски корма. В данном случае поведение определяется не присутствием внешнего раздражителя, а соответствующим внутренним побуждением.
· 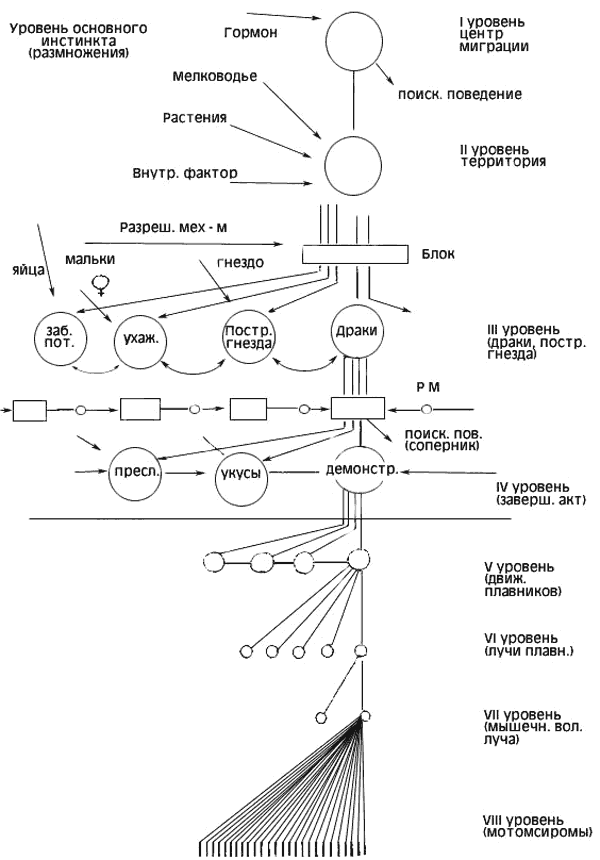
· Рис. 3. Схема организации инстинкта по Тинбергену (по: З.А. Зоринаб Л.В. Крушинский, Л.Г. Романова, И.И. Полетаева. Основные положения концепции К. Лоренца.)
· На уровне приобретенных форм поведения сложными последовательными поведенческими комплексами являются динамические стереотипы. Этот термин ввел И.П. Павлов, определивший "динамический стереотип" как
· устойчиво закрепившуюся систему условных нервных связей, обеспечивающую определенную интенсивность и последовательность реакций организма на воздействия внешней среды.
·
· Динамический стереотип возникает при достаточно длительном воздействии системы одних и тех же раздражителей (так называемом внешнем стереотипе). Он может существовать долгое время без какого-либо подкрепления. Предполагается, что физиологическую основу формирования начального этапа динамического стереотипа составляют условные рефлексы на время.
· Принципы координации рефлекторной деятельности
· Принцип сопряженного торможения или реципрокности -
· возникновение при возбуждении одного центра одновременного торможения другого центра для обеспечения цепных рефлексов и сложной функции.
· Пример реципрокности - регуляция спинным мозгом противоположных по функциональному назначению мышц конечностей. При возбуждении мотонейронов, иннервирующих мышцы-сгибатели правой ноги, реципрокно тормозятся мотонейроны мышц-разгибателей этой ноги, одновременно возбуждаются мотонейроны мышц-разгибателей левой ноги. Затем возникает возбуждение мотонейронов-разгибателей правой ноги и реципрокно — торможение мотонейронов-сгибателей правой ноги и возбуждение мотонейронов-сгибателей левой ноги. Таким образом, реципрокные взаимоотношения между указанными рефлексами обеспечивают цепной шагательный рефлекс.
· Реципрокные взаимоотношения есть и между рефлексами вдоха и выдоха, когда возбуждение центра вдоха тормозит центр выдоха и наоборот, что обеспечивает ритмичную смену фаз в процессе внешнего дыхания.
· 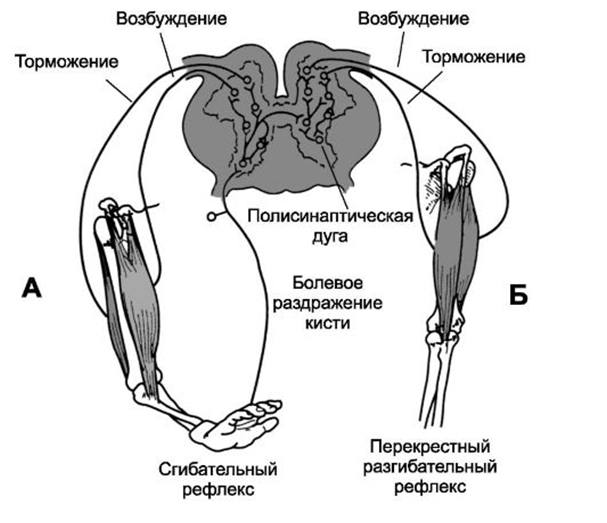
· Рисунок 1. Пример реципрокного торможения.
· Принцип доминанты (А.А. Ухтомский)
· - принцип деятельности нервной системы, проявляющийся в виде господствующей в течение определенного времени системы рефлексов, реализуемых доминирующими центрами, которые подчиняют себе или подавляют деятельность других нервных центров и рефлексов. Нейроны доминирующих центров приобретают более низкий уровень критической деполяризации мембран, т.е. становятся более возбудимыми, и способны эффективнее осуществлять пространственную и временную суммацию нервных импульсов. Синаптическое проведение к этим нейронам облегчено и поэтому они могут возбуждаться и за счет «посторонних» импульсов от не имеющих прямых связей с доминирующими центрами информационных каналов. Вследствие суммации многочисленных импульсов возбуждение отдельных нейронов и число возбужденных клеток в доминирующем центре нарастает и осуществляемые им рефлекторные реакции легко реализуются. Через систему вставочных нейронов доминирующий центр сопряженно тормозит другие центры и текущие рефлексы. Принцип доминанты позволяет концентрировать внимание и строить поведение для достижения определенной намеченной цели.
· Принцип общего конечного пути (Ч.Шеррингтон) -
· количественное преобладание чувствительных и других приходящих волокон над двигательными создает неизбежное столкновение импульсов в общем конечном пути, которым является группа мотонейронов и иннервируемые ими мышцы. Благодаря такому столкновению достигается блокирование всех воздействий, кроме одного, которое и регулирует протекание рефлекторной реакции; происходит селекция импульсов.
· Принцип обратной афферентации –
· заключается в рецепторном восприятии результатов рефлекторного акта и проведении информации назад в структуры нервного центра, где она обрабатывается и сравнивается с сохраняющимися параметрами возбуждения. Обратная афферентация реализуется в виде положительной или отрицательной обратной связи. Нервные центры осуществляют непрерывный контроль эффективности, целесообразности и оптимальности рефлекторной деятельности и при необходимости она корректируется.
· Принцип субординации нервных центров (принцип подчинения) -
· проявляется в виде регулирующего влияния выше расположенных нервных центров на ниже расположенные.
· См. выше описания экспериментов И.М. Сеченова (торможение центров спинного мозга как следствие возбуждения центров среднего мозга).
· Конфликты мотивации
· Рассогласования в поведении могут быть связаны с возникновением конфликтов, связанных с существованием разнонаправленной мотивации.
· К. Лоренц описал существование мотивационных конфликтов у животных, в случае, если одновременно возбуждаются два мотивационных центра.
· Классические примеры конфликтного поведения — поведение самцов территориальных видов, описанное Тинбергеном в результате наблюдений за трехиглой колюшкой и за различными видами чаек.
· Например, если самец А вторгается на территорию самца В, то последний нападает на него и преследует, а самец А спасается бегством. То же самое произойдет, если самец Б вторгнется на территорию самца А. Если же столкновение произойдет на границе этих двух территорий, то поведение обоих самцов будет выглядеть иначе: у обоих самцов элементы реакций нападения и бегства будут чередоваться. Причем элементы нападения будут выражены тем сильнее, чем ближе самец к центру своей территории. Напротив, по мере удаления от центра будут сильнее выражены элементы бегства.
· Такое смешанное поведение назвали "мозаичным".
· Как показали наблюдения за озерной чайкой , угрожающее поведение самцов на границе двух территорий включает пять поз, характер и последовательность которых зависят от реакций противника. Каждая из поз отражает определенную степень конфликта между противоположными внутренними побуждениями: агрессивностью — стремлением напасть на противника и страхом — стремлением убежать от него.
· Пример атласа поз серых гусей, отражающих разные соотношения между реакциями агрессии, страха и стремления приближения приведен на рисунке ниже.
· 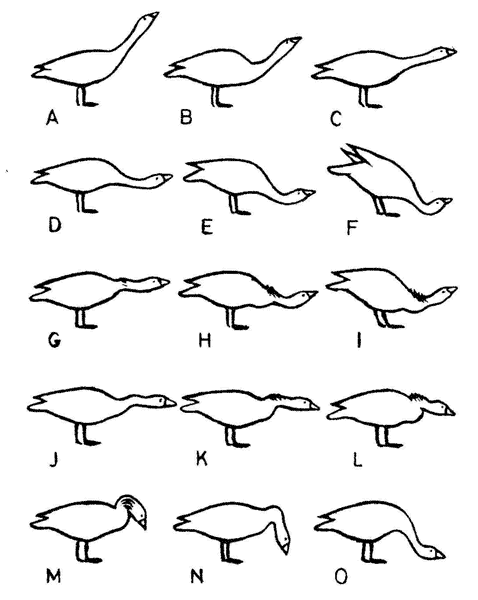
· Рис. 1. Атлас поз серого гуся. (По: К. Лоренц. Агрессия).
· Подобным образом с конфликтом мотивов связывают механизм так называемых «замещающих движений» (displacement activity), которые иногда также обнаруживаются у животных в конфликтных ситуациях. Например, в пограничной между двумя участками зоне два самца серебристой чайки (Larus argentatus), стоящие друг перед другом в угрожающих позах, могут внезапно начать чистить перья, белые гуси (Ansercoerulescens) на земле совершают те же движения, что и при купании, серые гуси (Anser anser) в этих ситуациях отряхиваются, а петухи клюют траву и все, что находится поблизости. Эти реакции являются врожденными, так как проявляются без соответствующего индивидуального опыта.
· Существуют два объяснения такого поведения.Первое - это переключение одной формы поведения на другую. Вторая - при "блокировке" равным возбуждением двух мотивационных центров проявляется третья, ранее заблокированная первыми двумя, доминирующая мотивация.
· (Подробнее см.: Мак-Фарленд Д. Поведение животных: Психобиология, этология и эволюция).
· Третья форма поведения, которая может проявиться при мотивационном конфликте - это смещенное поведение, при котором поведение реализуется, но в отношение иного объекта. Например, конфликт между социальным страхом и агрессией приводит к тому, что человек, разозлившись на начальника бьет по столу кулаком.
· Рис. 2. Примеры поведения, возникающего в результате конфликта мотиваций.
Дата: 2018-12-28, просмотров: 870.