1.Стадия прегенерализации.
Возникает выраженная концентрация возбуждения (главным образом, в проекционных зонах коры условного и безусловного раздражителей) и отсутствием условных поведенческих реакций.
2. Стадия генерализации.
В ее основе которой лежит процесс «диффузного» распространения (иррадиации) возбуждения. Условные реакции возникают на сигнальный и другие раздражители (явление афферентной генерализации), а также в интервалах между предъявлениями условного стимула (межсигнальные реакции). В этот период различные биоэлектрические сдвиги (блокада a-ритма, десинхронизация, θ-ритм и вызванные потенциалы) широко распространены по коре и подкорковым структурам. В период генерализации условного рефлекса наблюдается синхронизация биоэлектрической активности во многих участках коры и подкорковых образованиях – дистантная синхронизация биопотенциалов.
3.Стадия специализации.
Межсигнальные реакции угасают и условный ответ возникает только на сигнальный раздражитель. Изменения биотоков более ограничены и приурочены в основном к действию условного стимула. Этот процесс обеспечивает дифференцировку, тонкое различение стимулов, специализацию условно-рефлекторного навыка. В процессе специализации сфера распространения биопотенциалов значительно сужена и возрастает условно-рефлекторный ответ.
Процесс формирования условных рефлексов: теория Э.А. Асратяна
Если сигнальный и подкрепляющий раздражители вызывают собственные безусловные рефлексы, то они составляют нейросубстрат условного рефлекса. Действительно, условный раздражитель не является абсолютно индифферентным, так как сам вызывает определенную безусловно-рефлекторную реакцию - ориентировочную, а при значительной силе этот раздражитель вызывает безусловные висцеральные и соматические реакции. Дуга ориентировочного рефлекса также имеет многоэтажную структуру со своим корковым представительством.
Следовательно, при сочетании индифферентного раздражителя с безусловным (подкрепляющим) образуется временная связь между корковыми и подкорковыми ветвями двух безусловных рефлексов (ориентировочного и подкрепляющего), то есть образование условного рефлекса - это синтез двух или нескольких безусловных рефлексов.
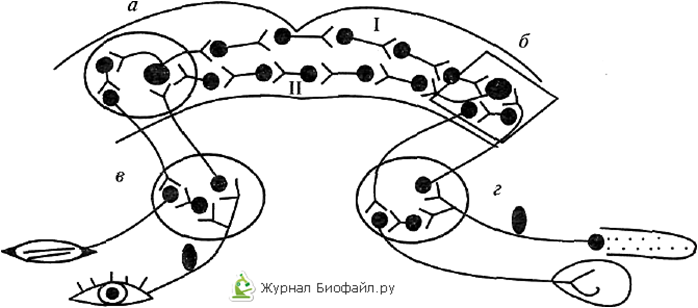
Рисунок 1.
Схема дуги условного рефлекса с двусторонней связью (по Э.А.Асратяну)
а — кортикальный центр мигательного рефлекса; 6 — кортикальный центр пищевого рефлекса; в, г — подкорковые центры мигательного и пищевого рефлексов соответственно; I — прямая временная связь; II — обратная временная связь.
Процесс формирования условных рефлексов по В.С. Русинову
В начальной стадии образования (генерализации) условный рефлекс представляет собой доминанту, а затем происходит дифференциация. Если с помощью прямой поляризации участка коры создать очаг возбуждения, то условно-рефлекторную реакцию можно вызвать любым индифферентным раздражителем.
Выделяется два механизма условно-рефлекторной деятельности:
надстроечный - регулирующий состояние мозга и создающий определенный уровень возбудимости и работоспособности нервных центров,
запускающий - который инициирует ту или иную условную реакцию.
Основные процессы, протекающие в нервной системе
Возбуждение и торможение
Возбуждение и торможение являются процессами, которые лежат в основе высшей нервной деятельности, в основе поведения. Взаимодействие этих процессов, их движение по коре полушарий и их последовательная смена позволяет осуществлять психические процессы. Все реакции организма обусловлены возбуждением определенных групп нервных клеток, а их прекращение - развитием торможения.
Возбуждение - это ответная реакция на раздражение, представляет собой биологический процесс, приводящий к изменениям обмена веществ и электрического потенциала клеточной мембраны и появлению биопотенциала.
Торможение - это процесс, выражающийся в предотвращении или угашении возбуждения.
Врожденные процессы нервной системы - безусловное возбуждение и безусловное торможение, на их основе в онтогенезе формируются условное возбуждение и условное торможение.
Сам процесс формирования условного рефлекса заключается в образовании связи между двумя очагами возбуждения, вызванными условным и безусловным раздражителями.
Доминанта - очаг возбуждения в центральной нервной системе, временно определяющий характер ответной реакции организма на внешние и внутренние раздражения. Доминирующий нервный центр (или группа центров) обладает повышенной возбудимостью и способностью стойко удерживать это состояние и тогда, когда первоначальный стимул уже не оказывает активирующего воздействия (инертность). Суммируя относительно слабые возбуждения др. центров, доминанта одновременно воздействует на них тормозящим образом. Основные положения учения о доминанте как общем принципе работы нервных центров сформулировал А. Ухтомский на основании выполненных им и его сотрудниками экспериментальных исследований (1911—1923).
Виды торможения
И.П. Павлов разделял все типы торможения на внешнее и внутреннее.
1. Внешнее торможение - реакция на внешний или внутренний стимул, затормаживающая условный рефлекс.
Любой посторонний раздражитель приводит к возникновению нового очага возбуждения в коре головного мозга, который ослабляет (или, в других случаях, усиливает) условно-рефлекторную деятельность. Новый очаг возбуждения, возникший в коре, является внешним по отношению к дуге выполняемого рефлекса. Внешнее торможение представляет собой врожденное безусловное свойство нервной системы.
Выделяют типы внешнего торможения:
· гаснущее, когда действующий раздражитель постепенно перестает вызывать ориентировочный рефлекс;
· неугасающее, возникающее при наличии какой-либо физиологической потребности или патологического процесса в организме;
· запредельное (охранительное) торможение, возникающее в нервной системе в ответ на очень сильные раздражители, когда работоспособность нервных клеток достигает предела, направлено на предохранение нервных клеткок от истощения.
2. Внутреннее (активное, условное) торможение. Наряду с образованием положительных условных рефлексов, в индивидуальной жизни животного образуется внутреннее торможение, служащее основой тормозных, или отрицательных, условных рефлексов. Такое торможение называют активным, или условным.
Выделяют виды условного торможения.
· Угасательное - в том случае, если условный раздражитель не сопровождается подкреплением, в результате этого постепенно теряет свое сигнальное значение, и рефлекс на него угасает.
Разные условные рефлексы без подкрепления угасают с неодинаковой скоростью. Более "молодые" и непрочные условные рефлексы угасают быстрее, чем "старые", прочные условно-рефлекторные связи. При угасании условного рефлекса происходит не просто разрыв условно-рефлекторной связи, а развивается активный тормозной процесс в коре головного мозга, который и подавляет условно-рефлекторную связь. Это положение подтверждается тем, что полностью угашенный условный рефлекс через некоторое время вновь восстанавливается.
Угасание условных рефлексов - биологически важное приспособление. Благодаря ему организм перестает напрасно тратить энергию - реагировать на сигнал, утративший свое значение. В силу данного обстоятельства, термин "условный рефлекс" в физиологии часто заменяется термином "временная связь".
· Дифференцировочное - развивается в коре головного мозга в том случае, если необходимо отличить один внешний раздражитель, являющийся условно-рефлекторным сигналом, от другого, сходного с ним раздражителя, который сигналом не является.
Дифференцировочное торможение участвует в образовании любого условного рефлекса, особо важно при формировании двух двигательных рефлексов на два различных раздражителя.
· Запаздывательное - при отсрочке подкрепления реакция оттормаживается и наступает в нужный момент. Позволяет экономить энергию.
Скорость образования внутреннего торможения зависит от индивидуальных особенностей ЦНС (соотношение процессов торможения и возбуждения, возрастные изменения и т.д.), от силы раздражителя (чем сильнее раздражитель, тем быстрее он становится тормозным).
Иногда постороннее раздражение, вызывающее сильную оборонительную реакцию, препятствует развитию внутреннего торможения и способствует проявлению угашенных условных рефлексов. Это явление называется растормаживанием.
На клеточном уровне выделяют следующие виды торможения.
I. По локализации.
1.Пресинаптическое торможение - осуществляется в пресинаптических элементах, связано с угнетением проведения нервных импульсов в аксональных окончаниях. Вставной тормозной аксон выделяет тормозной медиатор, который вызывает деполяризацию в постсинаптической мембране возбуждающего аксона. Возникшая деполяризация тормозит вход Са2 + из синаптической щели, приводит к снижению выброса возбуждающего медиатора в синаптическую щель, все это результирует в торможении реакции.
2.Постсинаптическое торможение - обусловлено выделением пресинаптическим окончанием аксона тормозных нейронов тормозного медиатора (ГАМК, глицин), который снижает или тормозит возбудимость мембран сомы и дендритов возбуждающего нейрона, с которым он контактирует, путем гиперполяризации мембраны.
Тормозные нейроны ЦНС – это специализированные вставочные нейроны, или интернейроны.
Виды тормозных нейронов:
1) клетки Реншоу;
2) клетки Уилкинсона;
3) грушевидные клетки Пуркинье (в мозжечке);
4) звездчатые клетки коры больших полушарий.
Некоторые медиаторы постсинаптического торможения:
1) гамма-аминомасляная кислота (ГАМК);
2) глицин;
3) норадреналин, дофамин, серотонин;
4) эндогенные бензодиазепины (эндозепины);
5) эндогенные опиоиды (эндорфины, энекефалины и др.), фактор J.
Дата: 2018-12-28, просмотров: 524.