| Показатели силы нервной системы | Суммарные индексы навязывания | |||||
| общий | дельта | тета | альфа | бета | гамма | |
| Абсолютный зрительный порог (24 испытуемых) | –409 ** | –253 | –390 * | –424 ** | –473 ** | –338 |
| ЭЭГ вариант угашения с подкреплением (19 испытуемых) | –470 ** | –220 | –275 | –562 ** | –451 * | –224 |
Примечание. р < 0,1; **р < 0,05.
Некоторым дополнением к данным настоящего раздела могут еще служить результаты эксперимента по определению эффекта навязывания как функции интенсивности стимула у детей. Этот эксперимент, поставленный К. Войку (1964), не имел целью сопоставление данных с показателями силы нервной системы, а поэтому мы не можем сказать точно, связаны ли индивидуальные особенности полученных функций с силой нервной системы испытуемых-детей. Однако весьма любопытно, что общая кривая зависимости эффекта навязывания от интенсивности стимула (рис. 66) очень близка к той асимптотической функции, которая была получена ранее для взрослых (рис. 54), а индивидуальные кривые демонстрируют примерно тот же характер индивидуальных различий, что и у взрослых. Это дает основания думать, что и у детей индивидуальные вариации проявления закона силы в реакции навязывания ритма, возможно, связаны с силой нервной системы (через связь этого параметра с абсолютной чувствительностью).
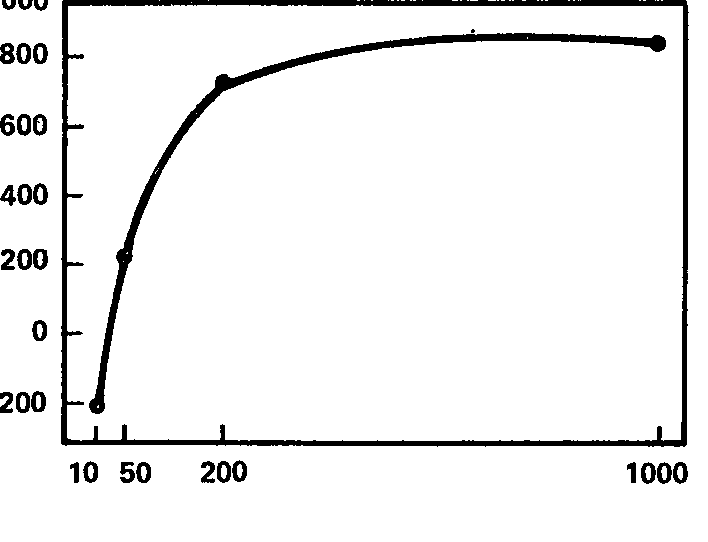
Рис. 66. Зависимость реакции навязывания ритма от интенсивности стимуляции у детей. Суммарная кривая. Ось абсцисс – интенсивность раздражения (в лк); ось ординат – ЭИНР (усл. ед.).
Что можно сказать по поводу материалов, изложенных в настоящей главе?
В трех различных экспериментах мы стремились получить опытные данные по вопросу о том, будут ли наблюдаться различия между сильными и слабыми нервными системами в количестве и динамике изменения исследуемого эффекта при возрастании стимуляции от каких-то минимальных значений (близких к порогу данной функции) до максимальных (находящихся на уровне предела данной физиологической функции). По-видимому, можно считать установленным, что эти различия действительно имеют место. Для слабых нервных систем характерны больший начальный эффект, более быстрое приближение к пределу и более раннее достижение предела данной функции, в то время как сильные нервные системы характеризуются, напротив, меньшим эффектом при минимальных значениях стимула, более медленным приближением к пределу функции и более поздним достижением этого предела. Для нас не существует иного объяснения этого явления, чем то, которое исходит из положения о взаимосвязи между силой нервной системы и абсолютной чувствительностью, можно полагать, что именно разница в уровне абсолютных порогов обусловливает указанные выше различия между сильными и слабыми нервными системами в количестве и динамике реактивного эффекта. Таким образом, через проверку следствий, вытекающих из закономерности связи между чувствительностью и силой, мы пришли к новому подтверждению самой этой закономерности.
Разумеется, каждый раз, как мы начинаем решать эту проблему с помощью конкретной методики, в дело вмешиваются специфические факторы этой методики, способные подчас сильно исказить ожидаемые «правильные» соотношения. В трех исследованных методиках такими интерферирующими факторами были (гипотетически) скорость проведения возбуждения по волокну в методике с временем реакции, лабильность периферических нервных элементов зрительного анализатора в методике КЧФ, лабильность корковых элементов зрительного анализатора в методике с навязыванием ритма. Устранить или обойти влияние этих специфических факторов при изучении обсуждаемой проблемы не всегда удается, но учитывать этот момент необходимо хотя бы для того, чтобы не делать поспешных отрицательных выводов.
В начале этой главы был поставлен частный вопрос относительно «параллельности», или «пропорциональности», прироста эффекта у сильных и слабых систем в промежуточной зоне стимуляции. Более или менее определенные данные для ответа на этот вопрос получены только в эксперименте с временем реакции, в той серии, где сопоставляются средние кривые «сильных» и «слабых» испытуемых при стимуляции в единицах порога, и, поскольку кривые фактически совпали, ответ на этот вопрос должен быть, по-видимому, положительным. Две другие примененные нами методики не дают такой легкой возможности для решения этой проблемы аналогичным образом. В эксперименте с фосфеном ввести стимуляцию в единицах порога чрезвычайно трудно, так как величина порога сама зависит от частоты стимуляции; поскольку опыт заключается как раз в плавном изменении частоты импульсов по мере нанесения раздражения, определить последнее в. единицах порога не представляется возможным (по крайней мере, без существенных технических усовершенствований аппаратуры). Что же касается навязывания ритма, то здесь для предъявления стимула в единицах порога следовало бы иметь прибор, представляющий собой комбинацию адаптометра со стробоскопом, а таким прибором мы не располагаем. Таким образом, от окончательного – положительного или отрицательного – ответа на вопрос о «пропорциональности» прироста эффекта в промежуточной зоне стимуляции следует пока воздержаться.
Дата: 2018-12-28, просмотров: 815.