| Номер испытуемого | Сохранение условной реакции к концу угашения с подкреплением, в % к исходной величине | Тип кривой |
| 1 | 193 | 1 |
| 2 | 182 | 1 |
| 3 | 145 | 1 |
| 4 | 143 | |
| 5 | 115 | 1 |
| 6 | 111 | 1 |
| Среднее | 131,5 | |
| 7 | 84 | 2 |
| 8 | 83 | 1 |
| 9 | 80 | 2 |
| 10 | 76 | 1 |
| 11 | 75 | 2 |
| 12 | 70 | 1 |
| 13 | 70 | 2 |
| Среднее | 77 | |
| 14 | 66 | 3 |
| 15 | 58 | 3 |
| 16 | 57 | 3 |
| 17 | 50 | 2 |
| 18 | 45 | 2 |
| 19 | 24 | 3 |
| 20 | 14 | 2 |
| Среднее | 45 |
1) из 8 кривых первого типа («пологих») 5 принадлежат испытуемым, у которых в результате угашения с подкреплением условная реакция увеличилась (первая группа), а другие 3 – испытуемым с маловыраженным падением условного эффекта (с сохранением 70–84 % первоначального уровня условной реакции – вторая группа);
2) из 7 кривых второго типа («крутых») 4 также принадлежат испытуемым второй группы, а 3 других – испытуемым с сильным снижением реакции к концу угашения с подкреплением (третья группа);
3) 4 кривых третьего типа (с «перегибом») принадлежат испытуемым последней, третьей группы;
4) наконец, одна «нетипичная» кривая (обозначается пунктиром на рис. 55) была получена у испытуемого первой группы.
Из сопоставления следует, что между силой нервной системы и особенностями динамики реакции навязывания ритма при возрастании интенсивности стимула существует достаточно отчетливая зависимость.
Конкретные примеры проявления этой зависимости даются на рис. 56–59, представляющих собой кривые записи реакций биотоков мозга в ответ на предъявление ритмических световых импульсов возрастающей интенсивности. На рис. 56 и 57 показана динамика реактивного эффекта испытуемых с сильной, а на рис. 58 и 59 – испытуемых со слабой нервной системой при частотах, соответствующих тета- и бета-ритмам ‑7 и 16 имп/с. Как электроэнцефалографические кривые, так и графики, построенные по результатам интегрирования, свидетельствуют о наличии подчас весьма резких различий между «сильными» и «слабыми» испытуемыми.
Для испытуемых с сильной нервной системой характерны в целом кривые с малым эффектом и малым приростом эффекта навязывания на протяжении всего диапазона интенсивностей. Испытуемым со слабой нервной системой свойственны кривые с несколько большим эффектом навязывания уже при минимальной интенсивности стимула и с заметным возрастанием этого эффекта по мере увеличения интенсивности; кроме того, некоторые из «слабых» кривых, видимо, отражают момент достижения предела и перехода за этот предел (в виде снижения эффекта).
Эти различия иллюстрируются рис. 60, на котором изображены средние кривые для групп «сильных» и «слабых» испытуемых.
Вычисление корреляционного отношения 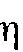 (эта), применяемого для измерения тесноты связи между двумя переменными в том случае, когда одна из них представляет собой градуальный ряд, а другая распределяется по нескольким качественным группам (P. Edwards, 1960), дает значение связи между кривой и силой нервной системы
(эта), применяемого для измерения тесноты связи между двумя переменными в том случае, когда одна из них представляет собой градуальный ряд, а другая распределяется по нескольким качественным группам (P. Edwards, 1960), дает значение связи между кривой и силой нервной системы 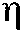 = 0,723 (р < 0,01). Таким образом, и статистический индикатор дает достаточно высокую меру связи между двумя сопоставляемыми характеристиками в том случае, когда для сравнения берутся кривые, усредненные по всем ритмам. Можно полагать, что на основе измерения ЭИНР как функции яркости мелькающего раздражителя может быть разработана достаточно «короткая» методика определения силы нервной системы. Мы проанализировали с этой целью кривые ЭИНР по отдельным физиологическим ритмам; для получения таких кривых обычно достаточно одного опыта с каждым испытуемым.
= 0,723 (р < 0,01). Таким образом, и статистический индикатор дает достаточно высокую меру связи между двумя сопоставляемыми характеристиками в том случае, когда для сравнения берутся кривые, усредненные по всем ритмам. Можно полагать, что на основе измерения ЭИНР как функции яркости мелькающего раздражителя может быть разработана достаточно «короткая» методика определения силы нервной системы. Мы проанализировали с этой целью кривые ЭИНР по отдельным физиологическим ритмам; для получения таких кривых обычно достаточно одного опыта с каждым испытуемым.


Рис. 56. (А, Б). Индивидуальная запись (исп. Щ., сильная нервная система). Стимуляция частотой 7 имп/с при различных уровнях интенсивности стимула не вызывает эффекта навязывания.
Обозначения: 1 – отметка раздражения, 2 – тета-ритм, 3 – альфа-ритм, 4 – бета-ритм, 5 – неанализированная ЭЭГ, 6 – отметка времени (1 с). Цифры справа обозначают результат интегрирования соответствующей частотной полосы (усл. ед.).

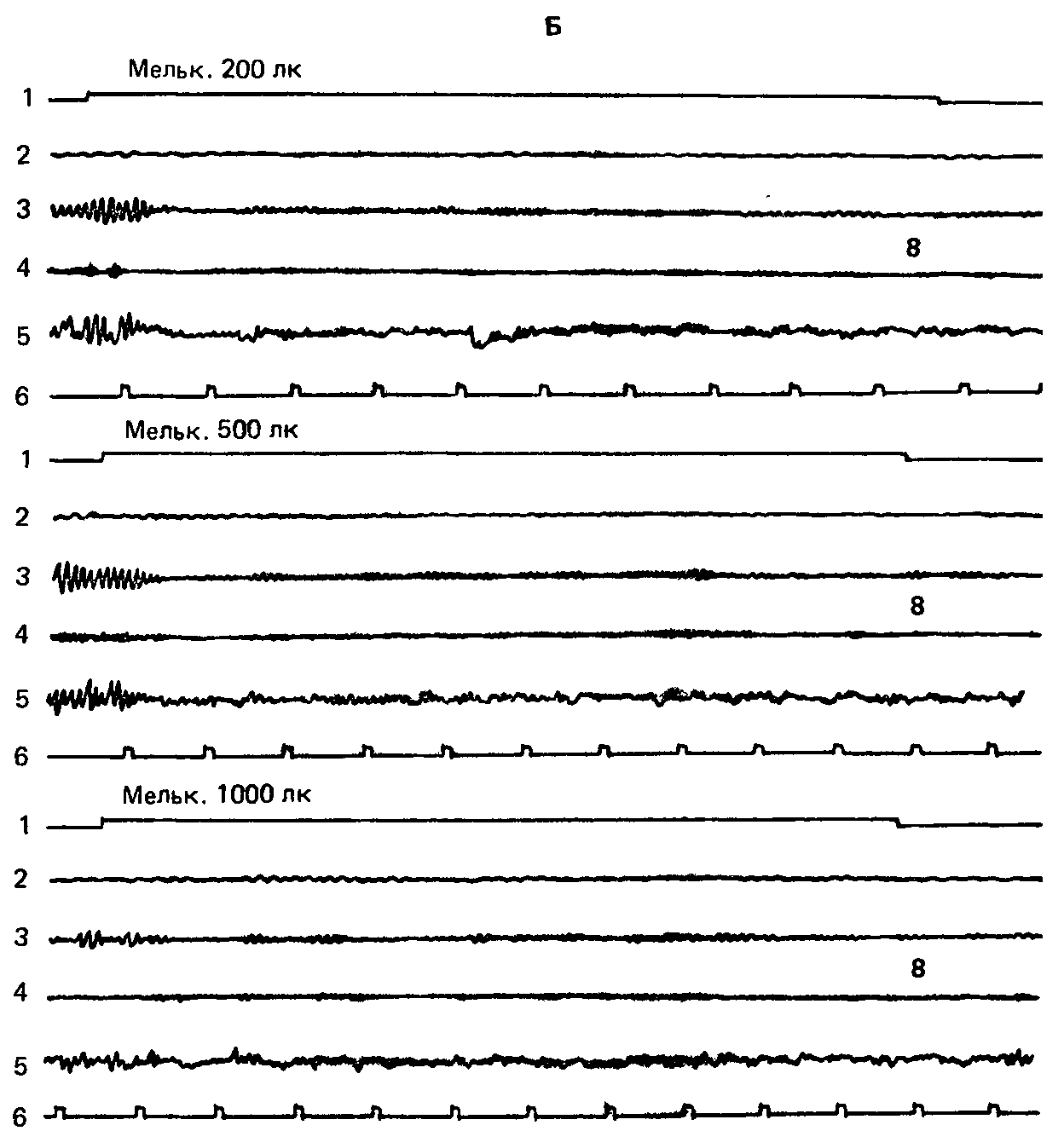
Рис. 57. (А, Б). Индивидуальная запись (исп. Щ., сильная нервная система). Стимуляция частотой 16 имп/с при различных уровнях интенсивности не вызывает эффекта навязывания. Обозначения те же, что на рис. 56.


Рис. 58. (А, Б). Индивидуальная запись (исп. К.О., слабая нервная система). Стимуляция частотой 7 имп/с вызывает отчетливо выраженную реакцию навязывания в тета-полосе, в целом возрастающую по мере увеличения интенсивности. Обозначения те же, что на рис. 56.


Рис. 59. (А, Б). Индивидуальная запись (исп. К.А., слабая нервная система). Стимуляция частотой 16 имп/с вызывает отчетливо выраженную реакцию навязывания в бета-полосе, возрастающую по мере увеличения интенсивности. Обозначения те же, что на рис. 56.
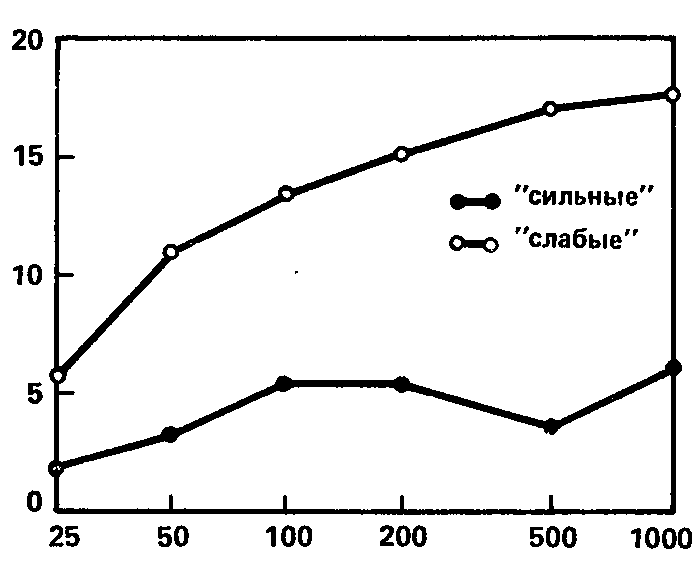
Рис. 60. Зависимость реакции навязывания от интенсивности раздражителя. Средние кривые для групп испытуемых с сильной и слабой нервной системой. Значения абсцисс и ординат те же, что на рис. 52 (В.Д. Небылицын, 1964).
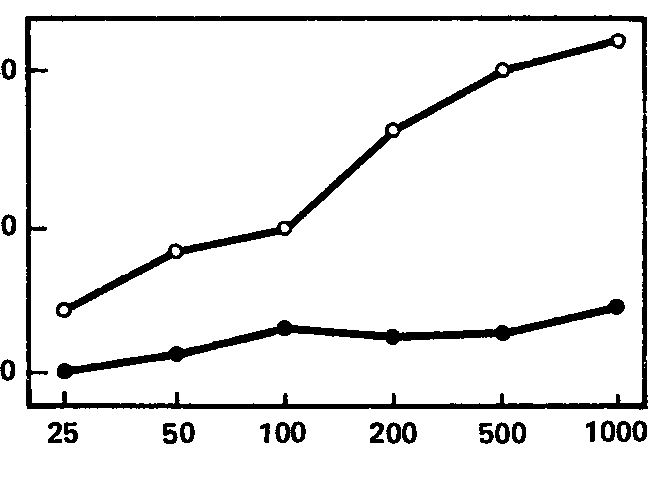
Рис. 61. Зависимость реакции навязывания ритма в тета-полосе от интенсивности раздражителя. Средние кривые для групп испытуемых с сильной (черные кружки) и слабой (белые кружки) нервной системой. Значения абсцисс и ординат те же, что на рис. 52.
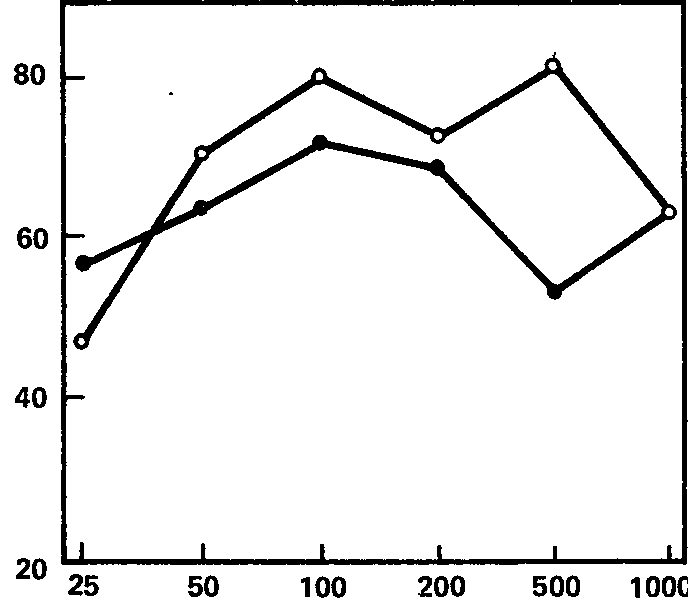
Рис. 62. Зависимость реакции навязывания ритма в альфа-полосе от интенсивности раздражителя. Средние кривые для групп с сильной и слабой нервной системой.
Обозначения те же, что на рис. 61.
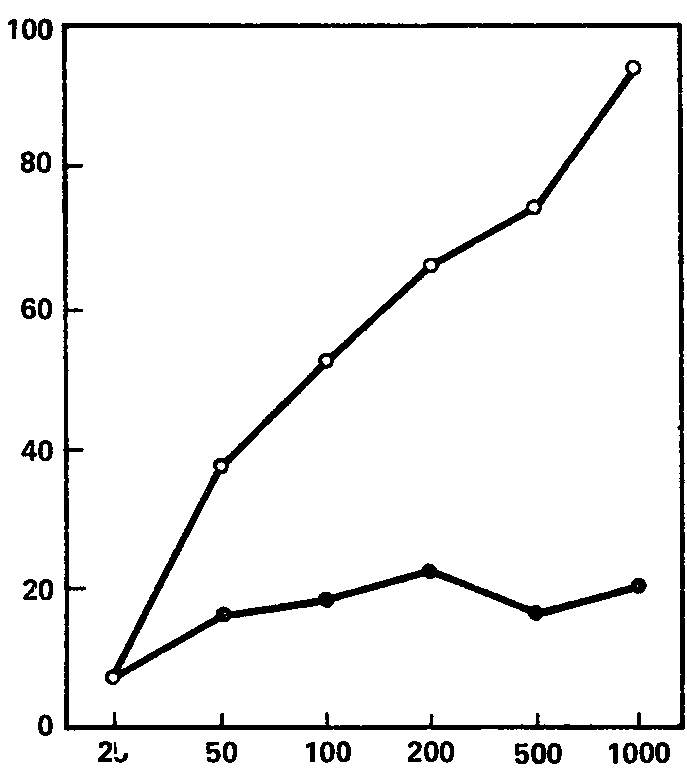
Рис. 63. Зависимость реакции навязывания ритма в бета-полосе от интенсивности раздражителя. Средние кривые для групп с сильной и слабой нервной системой.
Обозначения те же, что на рис. 61.
Эти кривые, для групп «сильных» и «слабых» испытуемых, показаны на рис. 61, 62, 63, из которых видно, что для частот, соответствующих тета- и особенно бета-ритму, кривые индивидов с сильной и слабой нервной системой существенно различны по своему характеру, особенно по степени наклона; правда, для диапазона альфа-ритма (рис. 62) различия между кривыми гораздо менее заметны. Отсюда вытекает, что для целей разработки короткой методики наибольшую эффективность обещает применение частот стимуляции, совпадающих с диапазоном тета- и бета-ритмов (по крайней мере, в пределах использованных нами частот). Причины же особого поведения диапазона альфа-ритма подлежат дальнейшему выяснению; возможно, они связаны со спецификой примененной методики оценки эффекта навязывания, тем более что другой метод оценки этой реакции, примененный Э.А. Голубевой, как мы увидим далее, дает при частотах полосы альфа-ритма довольно высокие величины связи с силой нервной системы.
Определенную перспективу в смысле разработки «короткой» методики обещает, как обнаруживается, и такой метод оценки реакции навязывания, как регистрация «кривых реактивности» по М.Н. Ливанову. Этот метод, состоящий в предъявлении индивиду ритмического светового раздражителя фиксированной частоты, но постепенно возрастающей яркости, удобен тем, что позволяет на протяжении одного «цуга» стимуляции, длящегося (в нашем случае) 38 с, наблюдать динамику реактивного эффекта на весь обычно применяемый диапазон интенсивностей светового раздражителя. При этом количественная оценка эффекта навязывания представляет определенные трудности, однако применение анализатора спектра позволяет все же выявить эту динамику с достаточной отчетливостью.
Опыт по определению индивидуальных особенностей «кривых реактивности» головного мозга и возможной зависимости их динамики от параметра силы нервной системы был поставлен нами с использованием в качестве источника световых импульсов постепенно возрастающей яркости фотостимулятора ЭФС‑01. От каждого из 25 испытуемых было получено 11 «кривых реактивности» – по числу примененных в опыте частот стимуляции, указанных в гл. V.
Сопоставление этих кривых с результатами угашения с подкреплением, основанное на визуальном анализе записей ЭЭГ составляющих и их изменений, позволяет, на наш взгляд, сделать вполне обоснованный вывод о существовании зависимости между силой нервной системы и проявлением эффекта навязывания в «кривых реактивности» коры больших полушарий.
Для испытуемых со слабой нервной системой оказалось характерным наличие эффекта навязывания почти на каждой из примененных частот стимуляции, кроме, может быть, минимальной ‑5 имп/с. При этом во многих случаях вершина эффекта наблюдалась не при максимальной яркости стимула, а при некоторых средних ее значениях. У испытуемых с сильной нервной системой реактивный эффект при данном способе стимуляции имел место гораздо реже, большей частью лишь в отдельных частотных диапазонах, а у некоторых из этих испытуемых он отсутствовал совершенно.
Мы не можем здесь за недостатком места сколько-нибудь широко иллюстрировать конкретными записями эти различия между «сильными» и «слабыми» испытуемыми. Приведем лишь для сравнения по одной характерной для каждой из групп записи биоэлектрической активности и ее изменений в ходе предъявления мелькающего света частотой 7 имп/с и постепенно возрастающей яркости. У «сильного» испытуемого (рис. 64) наблюдается только реакция блокады всех ритмов ЭЭГ на протяжении всего цуга стимуляции; у «слабого» (рис. 65) – отчетливо выраженная реакция навязывания первой гармоники, особенно при сильных значениях яркости стимула, а кроме того, временами и третьей гармоники (см. полосу бета-ритма).
Таким образом, при любом способе выявления эффекта навязывания обнаруживается зависимость динамики этой реакции в ходе изменения интенсивности стимула от параметра силы нервной системы.
И снова для объяснения природы этой зависимости нам приходится привлекать правило взаимосвязи между силой нервной системы и чувствительностью. Как и в опытах с временем реакции, а также с критической частотой фосфена, причиной различного хода «сильных» и «слабых» кривых при увеличении стимуляции, видимо, являются различия в абсолютной чувствительности сильных и слабых нервных систем. Большая чувствительность слабой нервной системы обусловливает больший эффект навязывания по сравнению с сильной нервной системой при минимальной интенсивности стимула (рис. 60) и более выраженный прирост этого эффекта («опережение») при увеличении стимула, с более стремительным приближением к пределу. Меньшая чувствительность сильной нервной системы и соответственно меньшая физиологическая эффективность стимуляции ведут к тому, что у «сильных» даже максимальная из применяемых интенсив
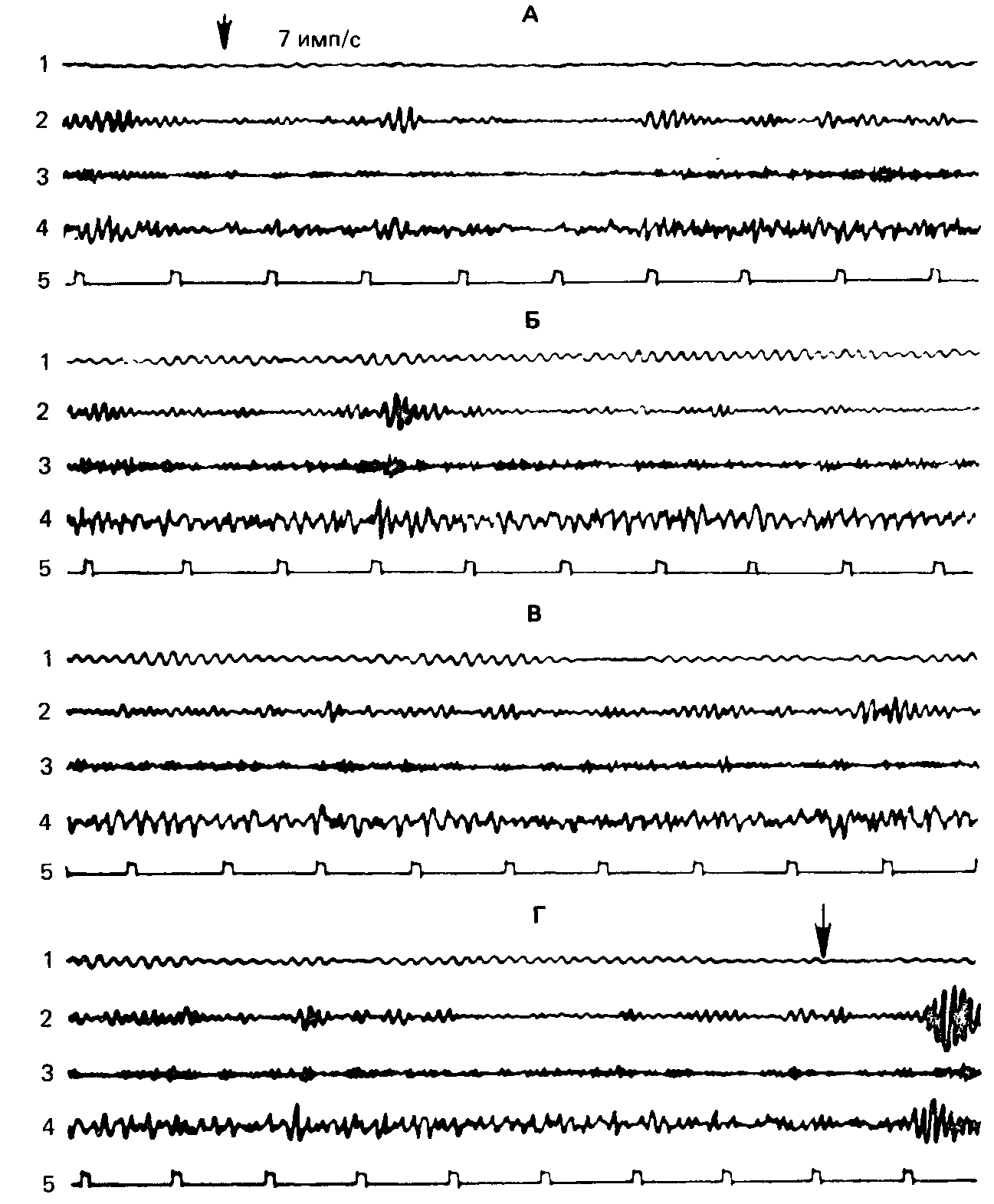
Рис. 64. Индивидуальная запись (исп. К., сильная нервная система). Отсутствие реакции навязывания ритма при стимуляции по методу «кривых реактивности», с частотой 7 имп/с.
Обозначения: А – начало, Б, В, Г – продолжение записи. 1 – тета-ритм, 2 – альфа-ритм, 3 – бета-ритм, 4 – неанализированная ЭЭГ, 5 – отметка времени (1с). Стрелками обозначены моменты включения и выключения стимуляции.
ностей стимула дает результат, лишь немного превышающий результат «слабых» при наименьшей яркости раздражителя (рис. 60). Таким образом, и эксперимент с навязыванием ритма, как это явствует из приведенных материалов, подтверждает следствия, вытекающие из правила взаимосвязи между чувствительностью и силой нервной системы, а следовательно, и само это правило.
К этому можно добавить, что прямой подсчет корреляции между суммарными индексами навязывания и абсолютными порогами, произведенный Э.А. Голубевой (1965), а также по ее данным в рамках работы по сопоставлению «коротких» методик (В.Д. Небылицын и др., 1965) приводит к довольно высоким, во многих случаях статистически значимым величинам коэффициентов (табл. 40), указывающим на то, что большая чувствительность зрительного анализатора способствует лучшему проявлению эффекта навязывания. Напомним, что методика оценки реакции навязывания, применяемая Э.А. Голубевой, значительно отличается от той, которая была использована нами. Тем более существенным кажется тот факт, что и при этой методике зависимость между показателями чувствительности (а также силы нервной системы – см. табл. 40) и индексами навязывания (которые в этой работе служат косвенными показателями подъема кривой, так как представляют собой сумму индексов для трех различных яркостей стимула) выявляется с достаточной отчетливостью.
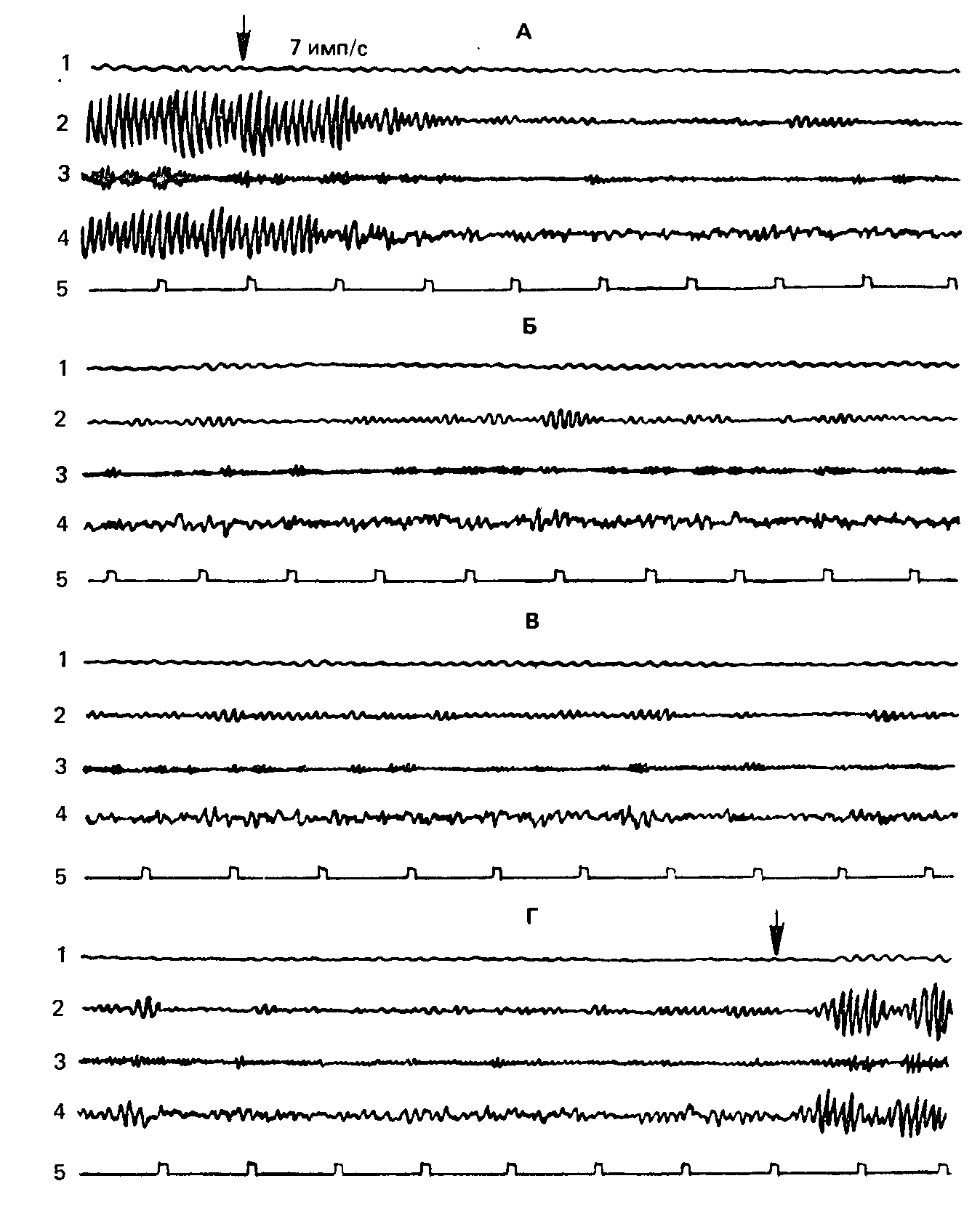
Рис. 65. Индивидуальная запись (исп. Ц., слабая нервная система). Отчетливо выраженная, имеющая максимум при средних значениях яркости стимула реакция навязывания ритма при стимуляции по методу «кривых реактивности», с частотой 7 имп/с.Обозначения те же, что на рис. 64.
Таблица 40
Дата: 2018-12-28, просмотров: 782.