| Пороги | Методики определения | силы | |||
| 1 | 2 | 3 | 4 | 5 | |
| Зрительные | 56** | 36* | 54** | 42* | 27 |
| Слуховые | 08 | 03 | 19 | 14 | 10 |
Примечания. 1) перечень силовых методик, обозначенных под номерами \ – 5, дается ниже; 2) • р < 0,05; >> р < 0,01.
В гл. 4 мы уже останавливались на индивидуальных особенностях изменений чувствительности при первых предъявлениях раздражителя – изменений, имеющих характер ориентировочной реакции и потому достаточно легко поддающихся угашению. Напомним, что эти изменения выражались у «сильных» испытуемых в снижении, а у «слабых» – в повышении чувствительности. Продолжение же стимуляции вызывало инверсию эффекта дополнительного стимула независимо от того, предъявлялся ли он кратковременными порциями, только на время каждого измерения порога, или действовал непрерывно в течение многих минут. Таким образом, после угашения ориентировочной реакции, которое обычно происходит очень быстро, дополнительный раздражитель вызывает повышение абсолютной чувствительности у лиц с сильной нервной системой и понижение абсолютной чувствительности у лиц со слабой нервной системой.
Описанная динамика может быть иллюстрирована рис. 42, на котором показаны для сравнения данные двух испытуемых, различающихся силой нервной системы. Эти различия, по Л.Б. Ермолаевой-Томиной, объясняются с физиологической точки зрения наличием у «сильных» испытуемых доминантного очага возбуждения в том анализаторе, к которому адресован пороговый раздражитель, и отсутствием такого очага у лиц со слабой нервной системой, для которых дополнительный стимул действует, следовательно, как внешний тормоз. Поскольку эти различия были достаточно определенны, методики, исследующие влияние прерывистого звука на зрительную чувствительность и прерывистого света на слуховую чувствительность, были включены в сопоставление ряда силовых методик (В.И. Рождественская и др., 1960), где показали удовлетворительные корреляции с референтными индикаторами силы нервной системы. Высокие корреляции этих методик с некоторыми вариантами индукционной методики были найдены также в разбираемой работе 3.Г.Туровской (19636). Это позволяет рассматривать индикаторы, основанные на определении характера отвлекающего действия дополнительного раздражителя, в качестве достаточно надежных показателей силы нервной системы.
Но вернемся к данным 3.Г.Туровской относительно связи между силой и чувствительностью.
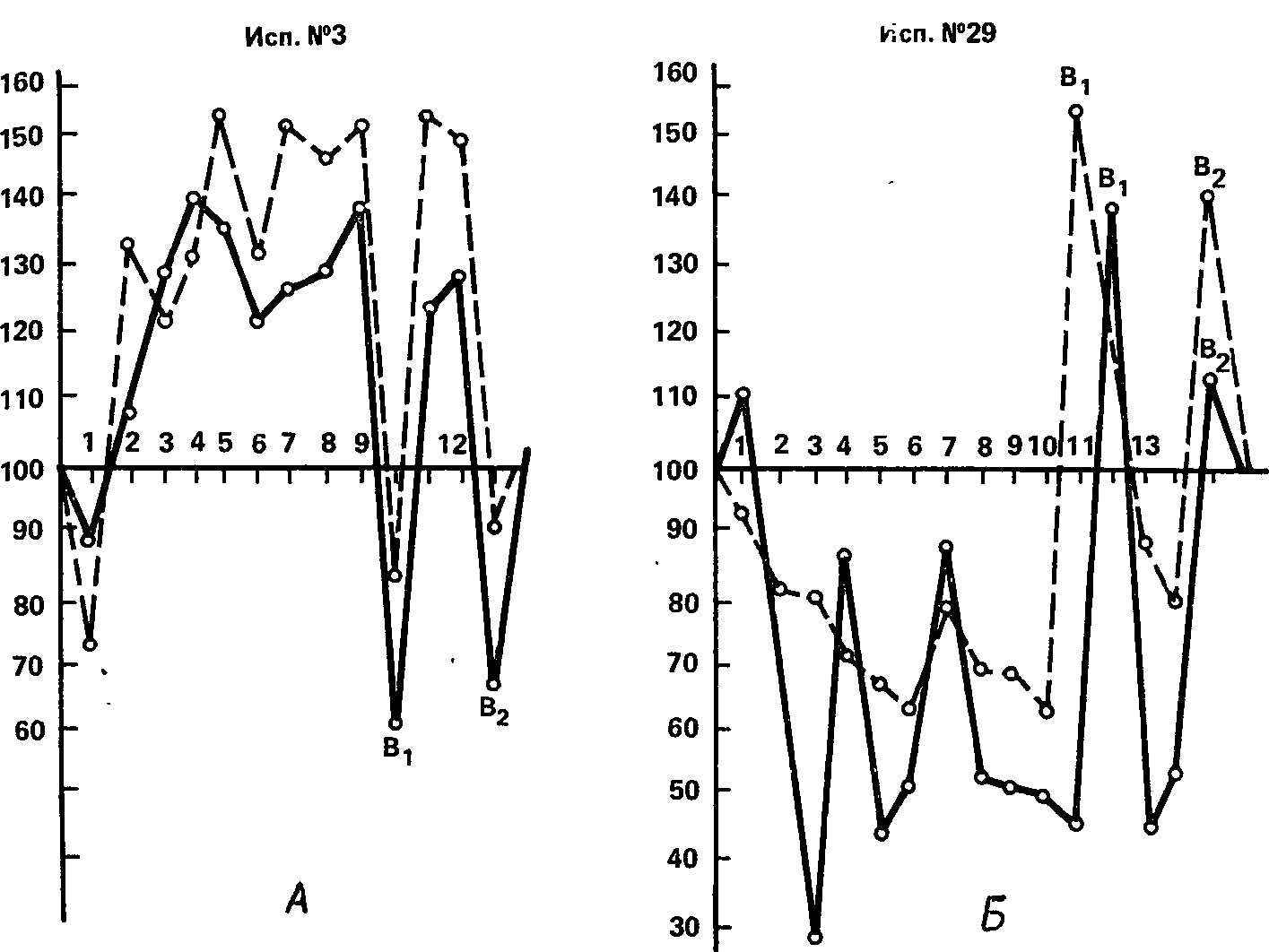
Рис. 42. Влияние длительно действующей гетеромодальной сенсорной стимуляции на абсолютную чувствительность испытуемых с сильной (А) и слабой (Б) нервной системой.
Сплошная линия – изменения абсолютной зрительной чувствительности при действии звука; штриховая линия – изменения абсолютной слуховой чувствительности при действии света. Bi и Во-уровни чувствительности при выключении дополнительного раздражения.
Ось абсцисс – порядковый номер изменений чувствительности в опыте; ось ординат – чувствительность (в %) но отношению к «фону» (Л.Б. Ермолаева-Томина, 1959).
Как явствует из перечня силовых методик, применяемых 3.Г.Туровской, три из них (1, 2 и 5) определенно относятся к зрительному анализатору; что касается двух других, то точная локализация сферы их действия представляется затруднительной, так как они имеюд дело одновременно с двумя анализаторами. Вероятно, этим и объясняется тот факт, что корреляции слуховых порогов с индикаторами силы хотя и положительны, но не достигают уровня значимости. В то же время зрительные пороги коррелируют с индикаторами силы нервной системы достаточно высоко, хотя и не во всех случаях.
Таким образом, с учетом ограничений, вводимых модальностью стимуляции, можно считать, что правило связи между чувствительностью и силой нервной системы, а говоря точнее, между зрительной чувствительностью и силой нервной системы в зрительном же анализаторе подтвердилось и в работе 3.Г.Туровской.
Таблица 26
Индивидуальные показатели слуховых порогов в сопоставлении с результатами ЭЭГ варианта угашения с подкреплением (В.Д. Небылицын, 19636)
| Испытуемые | Слуховой порог (в дБ от уровня 0,0002 бара) | Сохранение условной реакции в результате угашения с подкреплением (в % к исходной величине) | Испытуемые | Слуховой порог (в дБ от уровня 0,0002 бара) | Сохранение условной реакции в результате угашения с подкреплением (в % к исходной величине) | ||||
| Звук 70 дБ | Звук 90 дБ | Звук 70 дБ, кофеин 0,2 г | Звук 70 дБ | Звук 90 дБ | Звук 70 дБ, кофеин 0,2 г | ||||
| Р.А. | 23 | 97 | 94 | 108 | Л.Б. | 7 | 98 | 60 | 115 |
| Г.А. | 20,5 | 58 | 39 | 76 | П.В. | 6 | 27 | 31 | 18 |
| К.А. | 17,5 | 73 | 45 | 91 | М. | 5,5 | 31 | 13 | 90 |
| С.А. | 14,5 | 160 | 100 | 78 | Ш. | 5 | 46 | 27 | 140 |
| СБ. | 14 | 70 | 80 | 59 | Г.Б. | 4 | 60 | 78 | 35 |
| П.А. | 14 | 96 | 46 | 40 | П.Г. | 2,5 | 36 | 33 | 66 |
| к.в. | 13 | 54 | 26 | 95 | Г.В. | 2 | 44 | 53 | 80 |
| Р.Б. | 12 | 103 | 55 | 93 | Д. | 2 | 32 | 64 | 80 |
| Л.А. | 11 | 88 | 76 | 92 | к.г. | 1 | 39 | 38 | 38 |
| Р.В. | 10,5 | 52 | 63 | 63 | Средние | 9,7 | 68,7 | 56,5 | 76,4 |
| К.Б. | 10 | 82 | 70 | 49 | Стандартные отклонения | 5,93 | 31,6 | 24,2 | 28,1 |
| У. | 9 | 103 | 52 | 82 | |||||
| П.Б. | 8,5 | 62 | 102 | 92 | |||||
Что же касается слуховой чувствительности и силы в слуховом анализаторе, то подтверждение первоначальных данных о связи этих параметров было получено в работе с использованием ЭЭГ варианта угашения с подкреплением (В.Д. Небылицын, 19636). Здесь были применены три разновидности угашения с подкреплением: при использовании обычного звука – интенсивностью около 70 дБ от среднего порога, при использовании более громкого (на 20 дБ) звукового раздражителя и, наконец, с применением кофеина в дозе 0,2 г. Индивидуальные данные для каждого испытуемого по каждому из тестов приводятся в табл. 26.
Вычисление корреляции рангов между слуховыми порогами и первой разновидностью угашения с подкреплением дало значение р = 0,63 (р < 0,01), что означает для лиц с высокими порогами слуха статистически высокозначимую тенденцию к сохранению исходной величины условной рекции, а для лиц с низкими порогами – такую же тенденцию к падению условной реакции в результате угашения с подкреплением. Как видим, при совпадении сенсорных модальностей стимула, используемого для определения порога, и раздражителя, служащего условным сигналом в пробе на угашение с подкреплением, связь между чувствительностью и силой (собственно слабостью) выявляется достаточно отчетливо. Нужно, правда, заметить, что корреляции между порогами слуха и двумя другими модификациями угашения с подкреплением оказались гораздо ниже: для громкого звука р = 0,27 (р < 0,05) и для кофеина р = 0,20 (р >> 0,05). Однако этот факт легко поддается объяснению. Дело в том, что при громком звуковом раздражителе, как уже говорилось в предыдущей главе, имеет место увеличение эффекта угашения с подкреплением, затрагивающее главным образом индивидов с сильной нервной системой, так как у «слабых» испытуемых этот эффект – в виде близких к нулю условных реакций – достигается уже при обычном звуковом раздражителе. Это приводит к уменьшению диапазона индивидуальных различий, к подобию конечных результатов угашения с подкреплением «слабых» и «сильных» индивидов и, как следствие этого, к снижению величины коэффициента корреляции. Что же касается кофеина, то его применение, хотя и имеет, как уже указывалось выше, по существу противоположный эффект – повышение условных реакций, особенно у «слабых» испытуемых, приводит в конечном счете тоже к ликвидации различий между «сильными» и «слабыми» субъектами и к наблюдаемому уменьшению величины коэффициента корреляции. Таким образом, эти два коэффициента не противоречат общей картине достаточно высоких корреляций между чуствительностью и силой.
Последние (по времени) штрихи в эту картину внесены коллективным исследованием по сопоставлению ряда коротких методик определения свойств нервной системы (В.Д. Небылицын и др., 1965). Здесь абсолютная зрительная чувствительность была сопоставлена с ЭЭГ вариантом угашения с подкреплением, а также с индикаторами, которые в деталях будут описаны в следующей главе: с определением КЧФ, с наклоном кривой времени реакции как функции интенсивности звукового стимула и с временем реакции на слабые звуковые раздражители.
Результаты (табл. 27) были в определенном смысле парадоксальными, так как зрительные пороги коррелировали положительно, хотя значимо только в одном случае, с силовыми индикаторами, относящимися к слуховому анализатору, и не коррелировали с применявшимся ранее показателем методики КЧФ (суммой ординат кривой), адресующейся к зрительному анализатору. Однако первый факт не противоречит высказанному выше предположению о том, что зрительные пороги в силу самой природы зрительного анализатора фактически уже с самого начала, с рецепторного аппарата, являющегося частью центральной нервной системы, должны достаточно хорошо коррелировать с любыми адекватными индикаторами силы нервной системы, независимо от их модальности. Отсутствие же корреляции между зрительными порогами и суммой ординат кривой КЧФ, как мы полагаем, имеет своей причиной моменты главным образом методического характера. Если пренебречь этим несовпадением, то оказывается, что и в этом исследовании, особенностью которого была раздельная работа экспериментаторов, определяющих чувствительность и силу, и отсутствие в ходе работы взаимной информации о получаемых результатах, выявилось существование определенной связи между показателями чувствительности и индикаторами силы нервной системы.
Обозревая результаты цикла проведенных в лаборатории психофизиологии работ, в той или иной форме экспериментально исследовавших вопрос о связи между чувствительностью и силой, Б.М. Теплов отмечает: «Итак, сейчас надо говорить не о некоторой гипотезе, а об экспериментально доказанном на большом материале (в общей сложности более 150 испытуемых) закономерности обратной корреляции между силой нервной системы и чувствительностью» (1963, с. 24).
Таблица 27
Коэффициенты корреляции между зрительными порогами и некоторыми индикаторами силы нервной системы (В.Д. Небылицын и др., 1965)
| Угашение с подкреплением (ЭЭГ вариант) | Время реакции на очень слабый звук: средние данные двух опытов | Наклон кривой времени реакции | Сумма ординат кривой КЧФ | Разность второй и максимальной ординат кривой КЧФ | |
| Зрительные пороги | 472* | 300 | 319 | 091 | 273 |
Примечание. • р < 0,05.
До последнего времени экспериментальные факты в поддержку этой закономерности были получены только на человеке[32] при определении порога ощущения с помощью речевого отчета. Возможно, именно это давало некоторым полемически настроенным авторам повод для критики гипотезы, не взирая на последовательно получаемые факты. Тем более существенными представляются некоторые факты, полученные различными экспериментаторами на животных (собаках) и прямо свидетельствующие в поддержку закономерности, установленной на людях.
Так, М.В. Боброва (1960), сопоставляя характеристики реобазы и хронаксии мышечного аппарата собак с типологическими особенностями их, определенными по «малому стандарту», нашла совершенно четкую прямую зависимость между моторной реобазой (кстати, весьма тщательно измеренной) и предельной дозой кофеина, выдержанной животным. Иными словами, была обнаружена положительная корреляция между электрическим порогом возбуждения мышечной ткани и силой нервной системы, определенной «классическим» методом – по влиянию кофеина на рефлекторную деятельность. Минусом этой работы является малое число подопытных животных (четыре) и, следовательно, некоторая возможность случайного вывода.
Этого недостатка лишена другая работа, проведенная на 15 собаках и дающая, таким образом, вполне доказательный материал (Д.П. Неумывака-Капустник, А.И. Плаксин, 1964). Ее авторы провели детальное изучение показателей электровозбудимости нервно-мышечного аппарата в связи с типологическими особенностями нервной системы, в частности с ее силой. Сила нервной системы определялась при помощи кофеиновой пробы, суточного голодания и сверхсильных раздражителей. На основании этих испытаний авторы выделили 5 собак слабого и 10 сильного типа нервной системы. У всех животных была измерена реобаза разгибателя пальцев, а у части собак еще и реобаза других мышц.
Данные, полученные в этой работе, показаны в табл. 28, представляющей собой извлечение (с некоторой переработкой) из сводной таблицы, приводимой авторами. Сравнение величин убедительно свидетельствует о том, что порог раздражения у животных со слабой нервной системой в среднем значительно ниже, чем у «сильных» животных. К сожалению, авторы не воспользовались статистическими критериями для обоснования подмеченных соотношений, однако приводимый ими материал дает в одном случае возможность подсчитать коэффициент корреляции между силой нервной системы как качественным признаком (две группы – «сильных» и «слабых») и реобазой как количественным рядом (формула коэффициента корреляции для этого случая дается Эдвардсом – P. Edwards, 1960). Величина коэффициента оказывается равной 0,625 (р < 0,01); эта величина примерно того же порядка или даже выше, чем величины, обычно получаемые при работе с людьми.
Таблица 28
Средние величины реобазы мышечного аппарата у собак сильного и слабого типов нервной системы (Д.П. Неумывака-Капустник, А.И. Плаксин, 1964)
| Мышцы | Реобаза, в | |
| Сильный тип | Слабый тип | |
| М. extensor | 37,2(10) | 23,3(5) |
| М. flexor | 35,6(6) | 25,3(2) |
| М. auricularis | 41,2(6) | 24,8(2) |
Примечание. В скобках указано число подопытных животных.
Наконец, в подтверждение связи между чувствительностью и силой нервной системы можно сослаться на данные К.В. Кадарика (1962), который нашел, что если в группе беспородных собак 84 % (11 из 13) относились к сильному и только 16 % – к слабому типу нервной системы, то в группе чистопородных охотничьих гончих имеет место обратное соотношение и к слабому типу принадлежит 71 % собак (10 из 14). Автор в качестве возможного объяснения этого факта, ссылаясь на гипотезу Б.М. Теплова, выдвигает предположение о том, что отбор охотничьих собак по признаку развитого обоняния приводит тем самым к преобладанию среди них собак со слабой нервной системой. С этим, вероятно, можно согласиться, хотя прямые измерения порога обоняния автором не были произведены.
Итак, материалы целого цикла экспериментальных работ, проведенных на людях, существенно подкрепленные теперь группой фактов, полученных на животных, указывают на существование закономерной связи между абсолютной (не различительной!) чувствительностью и силой нервной системы. Эта связь выступает в виде положительного соотношения между силой нервной системы и порогами возбуждения сенсорной функции: при распределении контингента испытуемых по степени возрастания силы нервных клеток пороги возбуждения также будут иметь тенденцию к возрастанию (а чувствительность, возбудимость – к убыванию). Если это так, стало быть, соотношение между верхним порогом реакции возбудимой ткани – порогом запредельного торможения – и нижним порогом реакции – порогом возбуждения (ощущения, раздражения) – является относительно постоянным и может быть записано как 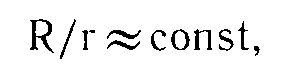 где R – верхний, а г – нижний порог реакции.
где R – верхний, а г – нижний порог реакции.
Это выражение означает, что диапазон между верхним и нижним порогами реакции в идеальном случае должен сохраняться от индивида к индивиду неизменным, но, конечно, в это соотношение в каждом отдельном случае вносятся определенные поправки, обусловленные влиянием факторов функционального порядка и приводящие к искажению соотношения, а в некоторых случаях, возможно, сводящие его на нет. Поэтому-то и можно говорить лишь об относительном постоянстве соотношения верхнего и нижнего порогов.
К сожалению, существующие методы оценки обоих порогов не дают возможности прямо сопоставить их величины, так как количественные характеристики верхнего и нижнего порогов даются в несоизмеримых единицах (результаты косвенной оценки этого соотношения изложены в следующей главе). Однако неоднократно наблюдавшиеся положительные коэффициенты корреляции между референтными показателями силы. нервной системы, из которых каждый дает приближенную оценку порога запредельного торможения, и абсолютными порогами, дающими меру чувствительности, ясно указывают на то, что хотя бы относительное, но все же постоянство соотношения между верхним и нижним порогами функции действительно существует.
Отсюда следует, что силу (выносливость) нервных клеток и их чувствительность можно рассматривать, по существу, как две стороны единого параметра жизнедеятельности нервного субстрата, связанного с интегральной, в высокой степени обобщенной функцией реагирования на интенсивность стимула. Это кардинальное свойство возбудимой ткани включает в себя как два неразрывно, связанных полюса одного и того же явления и чуткость системы к раздражению на его низшем – пороговом – уровне, и выносливость системы по отношению к воздействию на уровне предела ее функциональных возможностей.
Материал, изложенный в настоящей главе, позволяет подойти к решению часто обсуждаемого вопроса о биологическом смысле существования слабого типа нервной системы и причинах его появления в ходе естественной эволюции животного мира и человека. Мнения разных авторов о достоинствах и недостатках слабой нервной системы, о ее способности обеспечить нормальное «уравновешивание со средой» весьма различны. Как известно, И.П. Павлов в об-225 щем отрицательно оценивал возможности слабой нервной системы, называя ее «оранжерейной», «инвалидной» и т. д. Представление о «неполноценности» слабого типа нервной системы выражено в работах С.Н. Давиденкова (1947), Н.И. Красногорского (1954), Б.Н. Бирмана (1951) и других, о недостаточном «совершенстве» слабого типа – в статьях Р.Е. Кравецкого (1961), Н.Ф. Солодюк (1961) и других. Число подобных примеров можно было бы умножить. Однако, становясь на такую точку зрения, нелегко объяснить, почему слабый тип нервной системы не вымер давным-давно в процессе естественного отбора, в условиях конкуренции с «лучше приспособленными» особями сильного типа. Существование его в человеческой среде, а также в среде много веков назад прирученных домашних животных еще можно как-то объяснить, ссылаясь на отсутствие биологически обусловленной конкурентной борьбы в человеческом обществе и в условиях обитания домашних животных, хотя и здесь попытки объяснения встречают определенные трудности. Но тот факт, что индивиды слабого типа выявляются, скажем, среди обезьян, лишь недавно попавших в вольеры, или среди диких мышей и крыс, только что взятых для эксперимента, плохо поддается объяснению с «оценочной» позиции, занимаемой указанными авторами.
Другие исследователи придерживаются менее категорической точки зрения, допуская, что и слабый тип нервной системы обладает какими-то механизмами приспособления, обеспечивающими надлежащее уравновешивание со средой (Д.Р. Плецитый, 1957; Н.М. Вавилова и др., 1961; С.И. Вовк, 1961). Однако сущность этих компенсаторных или иных механизмов остается до сих пор не раскрытой.
Мы полагаем, что концепция, связывающая слабость нервной системы с более высокой чувствительностью, позволяет дать хотя бы частичный ответ на вопрос о биологической целесообразности существования слабого типа и механизмах его приспособления. Можно предположить, что именно высокая чувствительность животных слабого типа нервной системы, способность их уловить такие низкие по своей интенсивности сигналы, которые лежат ниже порога восприятия и, следовательно, ниже порога реакции особей сильного типа, и является той основой, на которой возможнее их соревнование с более выносливыми и в этом смысле действительно более приспособленными к жизни «сильными» индивидами.
В самом деле, более низкие сенсорные пороги означают возможность более ранней ориентировочной реакции при приближении врага или источника пищи. Они означают также возможность формирования условных реакций на такие интенсивности сигнала, которые еще не воспринимаются индивидами с более высокими порогами, и, вероятно, возможность ускоренного образования условных связей при физически равных интенсивностях стимула (в силу большей его эффективности для более чувствительной системы). Факты, говорящие в пользу последнего предположения, были получены в одной из наших работ, где оказалось, что у лиц с большей зрительной чувствительностью и соответственно со слабыми нервными клетками условные фотохимические реакции образуются значительно скорее, чем у испытуемых с противоположной характеристикой зрительного анализатора (В.Д. Небылицын, 19596). Аналогичные данные представлены Л.Б. Ермолаевой-Томиной (1963) на материале условных КГР, которые у «слабых» индивидов образовывались в среднем в 2 раза быстрее, чем у «сильных».
Иными словами, организация сенсорного аппарата слабой нервной системы такова, что она позволяет ее носителям во многих случаях избежать опасности, вместо того' чтобы столкнуться с ней лицом к лицу, разыскать пищу по малозаметным признакам, ускользающим от конкурентов, наконец, выработать систему реагирования и поведения, основанную на учете таких сигналов и признаков, которые недостаточны для более выносливых, более работоспособных, но менее чувствительных (а в известном смысле – и менее реактивных) особей, обладающих сильной нервной системой.
Можно полагать, что именно в этих особенностях слабой нервной системы заключается один из источников ее биологического преимущества, дающий ей возможность успешного соревнования в конкурентной борьбе в тех сферах жизнедеятельности, где на первый план выступают достоинства сенсорной организации.
Так, на конкретном примере взаимосвязи между слабостью и чувствительностью находит себе подтверждение общее правило о том, Что «каждое свойство нервной системы есть диалектическое единство противоположных с точки зрения жизненной ценности проявлений» (Б.М. Теплов, 1963, с. 25–26).
Дата: 2018-12-28, просмотров: 736.