Синтез фаговых белков. В первую очередь синтезируются ферменты, необходимые для образования копий фаговой ДНК. К ним относятся ДНК-полимераза, киназы (для образования нуклеозидтрифосфатов) и тимидилат синтетаза. Они появляются в клетке через 5-7 мин после её заражения. Клеточная РНК-полимераза транскрибирует вирусную ДНК в мРНК, которая транслируется бактериальными рибосомами в «ранние» белки фага, включая вирусную РНК-полимеразу и белки, способные посредством различных механизмов ограничивать экспрессию бактериальных генов. Вирусная РНК-полимераза осуществляет транскрипцию «поздних» белков (например, белков оболочки и эндолизина), необходимых для сборки фаговых частиц дочернего поколения. Некоторые вирусы расщепляют ДНК клетки-хозяина до нуклеотидов, чтобы использовать их для синтеза собственных нуклеиновых кислот.
Репликация нуклеиновых кислот реализуется за счёт активности вновь синтезированных вирусных ДНК-полимераз, производящих множественные копии вирусных нуклеиновых кислот.
Вновь синтезированные белки формируют в цитоплазме пул предшественников, входящих в состав головок и хвостов дочерних вирусных частиц. Другой пул содержит ДНК потомства. Специальные аффинные области в вирусной ДНК индуцируют объединение предшественников головок вокруг агрегатов нуклеиновой кислоты и образование ДНК-содержащих головок. Заполненная головка затем взаимодействует с хвостовой частью, образуя функциональный фаг. Весь процесс (от адсорбции до появления вновь синтезированных вирусов) занимает около 40 мин. После образования потомства клетка хозяина лизируется, высвобождая дочернюю популяцию. В разрушении клеточной стенки участвуют различные факторы: фаговый лизоцим, увеличенное внутриклеточное давление. Такой лизис бактериальной клетки называется лизисом изнутри (внутренний), но может быть и лизис извне (наружный) – происходит, когда на клетку адсорбируется сразу очень много бактериофагов, они проделывают многочисленные отверстия в клеточной стенке, через которые содержимое бактериальной клетки вытекает и она гибнет. В этом случае бактериофаг не размножается.
Если фаги вызывают лизис зараженной клетки с выходом в окружающую среду большого количества фаговых частиц, спрособных поражать новые клетки, то такие фаги называют вирулентными. В некоторых случаях вирулентных свойств фага оказывается недостаточно для разрушения бактерии. Подобные вирусы — умеренные фаги — претерпевают любопытные превращения, известные как редукция фага. При этом процессе ДНК вируса не вызывает образования вирусспецифических белков и нуклеиновых кислот, но включается в бактериальную хромосому. С практической точки зрения бактерия приобретает новый набор генов (встроенного вируса), а также становится «иммунной» к повторному заражению (интерференция вирусов). Подобный феномен известен как лизогения, а популяции бактерий — как лизогенные культуры. ДНК умеренного вируса реплицирует синхронно с размножением лизогенной бактерии, а иногда фаг начинает спонтанно размножаться, а клетка подвергается лизису. Некоторые умеренные фаги не способны образовывать дочерние популяции, то есть являются дефектными вирусами. Дефектные фаги используют как векторы в генной инженерии. Вирусная ДНК может длительно сохраняться в бактериальном потомстве. Такие латентные бактериофаги известны как провирусы, или профаги.
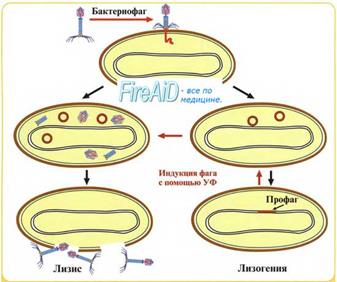 Пути развития умеренного фага.
Пути развития умеренного фага.
• Сохранение способности к инфицированию у умеренного фага зависит от низкомолекулярного белкового репрессора, кодируемого вирусной ДНК и «выключающего» все вирулентны функции бактериофага.
• Переход умеренного фага на литический цикл развития происходит при нарушениях синтеза белкового репрессора. При этом встроенный в геном бактерии вирус проявляет все свои вирулентные свойства, репродуцируется и лизирует клетки, а также может инфицировать другие бактерии. Переход умеренного бактериофага в литический цикл можно вызвать воздействием на бактерии ряда факторов. Например, если лизогенные культуры подвергнуть УФ-облученик действию или создать в среде избыток некоторых питательных веществ и витаминов, так происходит немедленная стимуляция вирулентных свойств фага — индукция фага.
Ассоциация фаговой ДНК с геномом бактерии способна качественно изменять свойства бактериальной клетки. Иными словами лизогенная бактерия будет вирулентной, тогда как её нелизогенный двойник останется безвредным. У других бактерий присутствие профага вызывает изменение морфологии или антигенных свойств. Такое изменение генетических свойств, вызванное вирусной ДНК, обозначают терми нами «инфекционная наследственность» или «лизогенная (фаговая) конверсия».
При лизогении происходит изменение наследственных свойств не только бактериальной клетки, но и фага; размножаясь в клетке, он способен захватывать некоторые гены бактерии и, инфицируя другую клетку, передаёт приобретённые гены новому хозяину. Подобная передача (трансдукция) во многом аналогична генетической рекомбинации у высших растений и животных.
Дата: 2018-12-28, просмотров: 417.