Бактериофаги [от бактерии, + греч. phagein, поедать] — группа вирусов, паразитирующих в бактериальных клетках.
История открытия и изучения фагов. В 1898 г. Н.Ф. Гамалея показал, что фильтрат сибиреязвенных бацилл вызывает лизис свежих культур этих микроорганизмов. 1915 г. Ф. Туорт обнаружил, что белые непрозрачные колонии стафилококков становились прозрачными и исчезали и что агент лизирующий стафилококки, проходит через бактериальные фильтры, сохраняя способность растворять свежие культуры этих микроорганизмов. Явление лизиса было описано, но природа не изучена. В 1917 г. канадский учёный д,Эрелль, изучая фильтраты испражнений больного дизентерией, которые он вносил в пробирки со свежезасеянной культурой возбудителя, заметил что при последовательных посевах после инкубации их в термостате лизирующая способность фильтрата возрастает до полного растворения дизентерий ной культуры ( это совпало с началом выздоровления больного). Из этого учёный сделал вывод, что растворяет её живой агент, проходящий через бактериальный фильтр, т.е. вирус. Открытый вирус д,Эрелль назвал бакрериофагом , а явление – бактериофагией.
Вирусы, вызывающие гибель инфицированных бактерий, известны как литические бактериофаги . Размножение и выход дочерних популяций вируса из бактерии сопровождается её гибелью и разрушением (лизисом).
Бактериофаги устойчивы к различным физическим и химическим воздействиям. Большинство из них без вреда переносит высокие температуры (50-70 °С), действие дезинфектантов (за исключением кислот и формалина), прямой солнечный свет и УФ-облучение в низких дозах. Бактериофаги проявляют иммуногенные свойства, вызывая синтез специфических антител. Бактериофаги широко распространены в природе — их выделяют из воды, почвы, организмов различных животных и человека.
Принципы классификации бактериофагов аналогичны подходам к систематике вирусов вообще. В основу классификации положены антигенная структура, морфология фагов, спектр действия, химический состав и др. Большинство фагов относится к ДНК-содержащим вирусам с нуклеокапсидом, организованным по принципу смешанной симметрии.
Именуют фаги обычно по названию клкетки-хозяина (дизентерийный, стрептококковый и т.д.), по группе хозяина фаги могут и классифицироваться. Фаги обладают строгой специфичностью. По спектру действия (по специфичности) выделяют типовые фаги (Т-фаги), лизирующие бактерии отдельных типов внутри вида, моновалентные (видовая специфичность) фаги, лизирующие бактерии одного вида, и поливалентные фаги, лизирующие бактерии нескольких видов.
Строение бактериофагов наиболее полно охарактеризовано на основе изучения Т-фагов кишечной палочки. Внешне большинство бактериофагов напоминают сперматозоиды или головастиков, но среди них встречают и другие формы, на основании которых выделяют пять основных типов бактериофагов.
• К типу I бактериофагов относят ДНК-содержащие нитевидные фаги, лизирующие бактерии, содержащие F-плазмиды.
• Фаги типа II представлены головкой и рудиментом хвоста. Геном большинства из них образован молекулой РНК.
• Бактериофаги типа III имеют короткий хвост.
• К типу IV относят фаги с несокращающимся хвостом и двухнитевой ДНК .
• Фаги типа V имеют ДНК-геном, сокращающийся чехол хвоста, который заканчивается базальной пластиной.
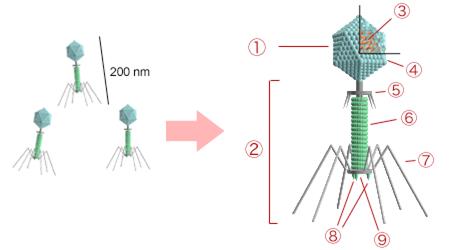
Рис.5 Строение бактериофага. 1 - Головка, 2 - хвостик, 3 - ДНК, 4 - капсид, 5 - воротник, 6 - чехол, 7 - хвостовые фибриллы, 8 - шипы, 9 - базальная пластинка.
Головка Т-фагов ( бактериофагов ) образована из однотипных субъединиц, организованных по принципу кубической симметрии, и может достигать размеров 100 нм. Капсомеры головки состоят из белковых молекул, построенных преимущественно из аспарагиновой и глутаминовой кислот, а также лизина. Содержание белка и ДНК в головке примерно одинаково. Геном большинства фагов образует спирально упакованная двойная нить ДНК. В составе фаговой ДНК обнаружены необычные азотистые основания (например, оксиметилцитозин).
Хвост Т-фагов ( бактериофагов ) может достигать 250 нм в длину и 25 нм в ширину. Он включает поль - стержень (сконструирован по принципу спиральной симметрии) и сократительный чехол, присоединяющийся к воротничку, окружающему стержень около головки. Чехол образован 120-140 белковыми молекулами, каждая из которых связывает одну молекулу АТФ и ионы Са2+. В дистальном отделе стержня расположена шестиугольная базальная пластина с шестью шипами, шестью нитями (фибриллами). У чётных фагов окончания фибрилл опущены вниз, а у нечётных — загнуты вверх. У некоторых Т-фагов ( бактериофагов ) в дистальной части хвоста находится лизоцим (эндолизин).
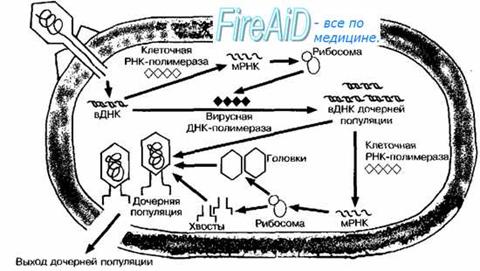
Взаимодействие бактериофагов с клеткой специфично, так как они, как правило, инфицируют бактерии только определённого вида. Подобно вирусам животных, репродуктивный цикл литических бактериофагов включает адсорбцию свободного фага на клетке, инъекцию ДНК, репродукцию фага, выход дочерних популяций.
Прикрепление фага к бактерии (адсорбция) происходит при помощи поверхностных структур бактериальной стенки, служащих рецепторами для вирусов (в липополисахаридном слое, в наружной мембране). На бактериях без клеточной стенки (протопласты, L-формы) бактериофаги не адсорбируются. Некоторые фаги в качестве рецепторов используют F-пили. Помимо рецепторов, адсорбция фага зависит от рН среды, температуры, наличия катионов и некоторых соединений. При избытке фага на одной клетке может адсорбироваться до 200-300 вирусных частиц.
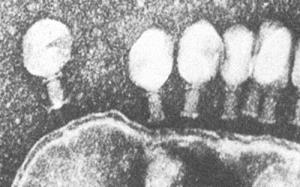

После адсорбции происходит ферментативное расщепление клеточной стенки лизоцимом, находящимся в дистальной части отростка. Базальная пластина хвоста лизирует прилегающий фрагмент клеточной стенки, выделяя присутствующий в отростке лизоцим. Одновременно в чехле высвобождаются ионы Са2+, активизирующие АТФазу, что вызывает сокращение чехла и вталкивание стержня хвоста через ЦПМ в клетку. Затем вирусная ДНК впрыскивается в цитоплазму (внедрение вирусной ДНК). Поскольку диаметр канала лишь немного превышает диаметр молекулы ДНК (около 20 нм), то ДНК способна попадать в цитоплазму только в форме нити.
Проникнув в клетку, ДНК фага «исчезает»; уже через несколько минут обнаружить вирус не удаётся. В этот, так называемый скрытый период (эклипс) вирус берёт на себя генетическое управление клеткой, осуществляя полный цикл репродукции фага. К его окончанию составляющие фага соединяются в зрелый вирион.
Дата: 2018-12-28, просмотров: 419.