Невозможно непосредственно определить обмен энергии в мышечных волокнах. Множество косвенных лабораторных методов позволяют определить интенсивность и расход энергии в покое и при выполнении физической нагрузки. Многие из этих методов применяются с начала XX ст. Другие лишь совсем недавно стали использоваться в физиологии мышечной деятельности. В следующих параграфах мы рассмотрим некоторые из этих методов.
94
ПРЯМАЯ КАЛОРИМЕТРИЯ
Как уже отмечалось, только около 40 % энергии, высвобождающейся при метаболизме глюкозы и жиров, идет на образование АТФ. Остальные 60 % превращаются в тепло, поэтому интенсивность и количество освобождаемой энергии можно определить, измерив образуемое тепло. Этот метод называется прямой калориметрией.
Впервые он был описан Зунцом и Хагеман-ном в конце XIX ст. Они создали калориметр (рис. 5.10), представляющий собой герметически закрытую камеру. Стенки камеры имеют медные трубы, по которым течет вода. Когда человек находится в камере, тепло, выделяемое его телом, испаряется и нагревает воду. Регистрируются изменения температуры воды, а также воздуха, поступающего в камеру и выходящего из нее во время дыхания человека. Эти изменения обусловлены теплом, выделяемым телом человека. Используя суммарные показатели, можно количественно оценить метаболизм.
Калориметры очень дороги, кроме того, для получения достоверных результатов период исследования должен длиться довольно долго. Единственным реальным преимуществом калориметров является то, что они непосредственно измеряют тепло. Хотя калориметр позволяет точно
определить общий расход энергии, тем не менее он не дает возможности проследить за быстрыми изменениями в высвобождении энергии. Именно поэтому энергетический обмен во время интенсивной нагрузки нельзя изучать с помощью калориметра. Метод прямой калориметрии довольно редко используется в наше время. Гораздо проще и дешевле определить расход энергии на основании обмена кислорода и диоксида углерода во время окислительного фосфорилирования.
НЕПРЯМАЯ КАЛОРИМЕТРИЯ
Как уже отмечалось, метаболизм глюкозы и жиров зависит от наличия О-, образования СОд и воды. Количество обмениваемых Од и СОд в легких обычно равно тому их количеству, которое используется и выделяется тканями тела. Зная это, можно определить расход энергии, измерив количество дыхательных газов. Этот метод называется непрямой калориметрией, поскольку образование тепла измеряется косвенно, на основании дыхательного газообмена СО^ и Од.
Прибор для измерения образования СО^ и потребления О- показан на рис. 5.11. Несмотря на то, что он несколько громоздок и ограничивает движения, его можно использовать в различных условиях в лаборатории, на игровой площадке и т.д.
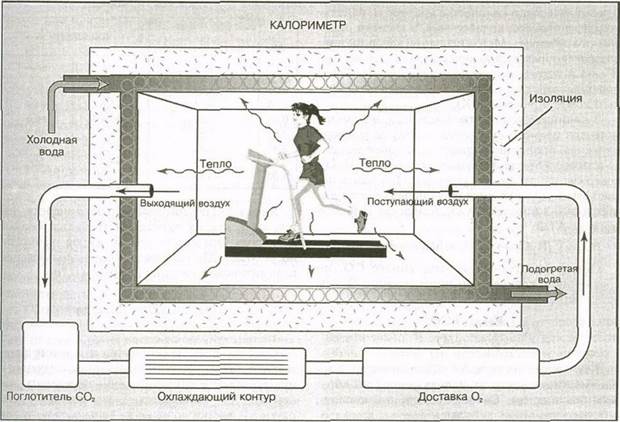
Рис. 5.10. Калориметрическая камера 95
| ДК= V co2| V o2 |
| бы полностью окислить эту молекулу до СО, и НдО, необходимо 23 молекулы кислорода: 16С + 160, -> 16СО, 32Н + 8o2 -16Н;0 Всего необходимо 24 o2 — 1 o2 обеспеченная самой пальмитиновой кислотой 23 o2 должны быть добавлены В конечном итоге этот процесс окисления приводит к образованию 16 молекул С o2 16 молекул Н2o и 129 молекул АТФ: |
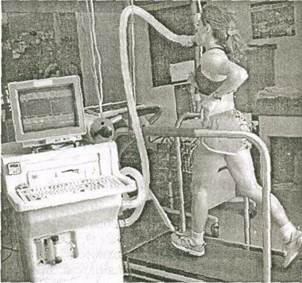
Рис. 5.11. Приборы, используемые для измерения потребления кислорода и образования СО;
Дыхательный коэффициент
Для определения количества энергии, расходуемой организмом, необходимо знать, что используется в окислительных процессах в качестве субстратов (углеводы, жиры или белки). Содержание углерода и кислорода в глюкозе, свободных жирных кислотах и аминокислотах очень отличается. В результате этого количество кислорода, используемого в процессе метаболизма, зависит от окисляемого субстрата. Метод непрямой калориметрии позволяет определить количество выделяемого СО^ (Усо.) и потребляемого кислорода ((^со,). Соотношение этих двух величин называется дыхательным коэффициентом (ДК).
В принципе количество кислорода, необходимого для полного окисления молекулы углевода или жира, пропорционально содержанию углерода в этом субстрате. Например, глюкоза (С^Н,,Од) содержит шесть атомов углерода. При сжигании глюкозы 6 молекул кислорода используются для образования 6 молекул СО;, 6 молекул Н^О и 38 молекул АТФ:
60, + С6h12o6, -> 6СО, + 6Н,0 + 38АТФ.
Оценивая количество выделенного СО, по сравнению с количеством потребленного О,, мы установим, что ДК равен 1,0:
ДК=
| 6СО |
| 60. |
| ^=1,0 |
Как следует из табл. 5.4, ДК изменяется в зависимости от субстрата, используемого для образования энергии. Свободные жирные кислоты содержат значительно больше углерода и водорода, но меньше кислорода, чем глюкоза. Возьмем, например, пальмитиновую кислоту, С^Н^Оу Что-
Для сжигания одной молекулы жира требуется значительно больше кислорода, чем для сжигания одной молекулы углеводов. Во время окисления углеводов из каждой молекулы используемого О-, образуется приблизительно 6,3 молекул АТФ (38 АТФ на 6 О-), тогда как при обмене пальмитиновой кислоты — 5,6 молекул АТФ (129 АТФ на 23 О;).
Таблица 5.4. Калорическая эквивалентность ДК и количество килокалорий за счет окисления углеводов и жиров
| Килокалории,%
| |||
| ДК | Энергия, ккал-л ' 0; |
| |
| углеводы | жиры | ||
| 0,71 | 4,69 | 0 | 100 |
| 0,75 | 4,74 | 15,6 | 84,4 |
| 0,80 | 4,80 | 33,4 | 66,6 |
| 0,85 | 4,86 | 50,7 | 49,3 |
| 0,90 | 4,92 | 67,5 | 32,5 |
| 0,95 | 4,99 | 84,0 | 16,0 |
| 1,00 | 5,05 | 100,0 | 0 |
Хотя жиры обеспечивают больше энергии, чем углеводы, для их окисления требуется больше кислорода. Это значит, что ДК жиров значительно уступает ДК углеводов. При использовании палимитиновой кислоты ДК равен 0,70:
Определив ДК на основании измерения дыхательных газов, сравним полученное значение с приведенным в табл. 5.4, чтобы определить, какой субстрат окисляется. Если, например, ДК равен 1,0, клетки используют только глюкозу или гликоген, и каждый литр потребляемого кислорода образует 5.05 ккал энергии. При окислении
96
| 97 |
одних только жиров образуется 4,69 ккал-л^О^, а
при окислении белков — 4,46 ккал-л'1 потребленного Оу
Следовательно, если мышцы используют только глюкозу, а организм потребляет 2 л кислорода в минуту, интенсивность теплового образования энергии будет 10,1 ккал-мин'' (2 л-мин''-5,05 ккал).
Ограничения
Даже метод непрямой калориметрии далек от совершенства. Измерения газообмена предполагают, что содержание О, в организме остается постоянным и что обмен СО- в легких пропорционален количеству его, выделяемому из клеток. Артериальная кровь остается почти полностью насыщенной кислородом (около 98 %) даже при интенсивном усилии. Мы можем предположить, что количество кислорода, извлекаемого из воздуха, которым мы дышим, пропорционально его количеству, потребляемому клетками. Отметим, что обмен диоксида углерода менее постоянен. Депо СО^ в организме довольно большие; их можно изменить либо глубоким дыханием, либо выполнением физического упражнения высокой интенсивности. При таких условиях количество СОу выделяемого в легкие, может не соответствовать производимому в тканях. Таким образом, определение количества используемых углеводов и жиров на основании измерений газов можно считать достаточно надежным только в состоянии покоя или при выполнении упражнений определенной интенсивности.
Использование дыхательного коэффициента также может привести к неточностям. Вспомним, что белок окисляется в организме не полностью, поскольку азот не способен окисляться. Поэтому на основании ДК невозможно определить величину использования белков. Поэтому ДК иногда называют небелковым ДК, поскольку он не учитывает окисление белков.
Традиционно считалось, что вклад белков в образование энергии во время физической нагрузки незначителен, поэтому физиологи при проведении подсчетов использовали небелковый ДК. Однако результаты последних исследований показывают, что во время физической нагрузки продолжительностью несколько часов вклад белков в образование энергии может составлять до 10 %.
Организм, как правило, использует сочетание источников энергии. ДК изменяется в зависимости от своеобразия окисляемого субстрата. В покое ДК обычно составляет 0,78 — 0,80. Во время нагрузки мышцы в основном используют углеводы в качестве источника энергии, что ведет к повышению ДК. С увеличением интенсивности нагрузки возрастает потребность в углеводах. По мере увеличения количества используемых углеводов ДК стремится к 1,0.
7,-„
Это приближение ДК к 1,0 отражает потребность в глюкозе крови и мышечном гликогене, а также может свидетельствовать о том, что значительно большее количество СО^ выделяется из крови, чем производится мышцами. По мере наступления состояния изнеможения в крови аккумулируется лактат. Организм пытается реверсировать это окисление, выделяя большее количество СО^. Накопление лактата увеличивает образование СО-, поскольку избыток кислоты ведет к превращению угольной кислоты крови в СО^. Вследствие этого избыток СО^ диффундирует из крови в легкие, увеличивая количество выделяемого СОу Поэтому ДК, достигающий 1,0, может не совсем точно определять тип субстрата окисления, используемого мышцами. Кроме того, образование глюкозы вследствие катаболизма аминокислот дает ДК ниже 0,70. Следовательно, определение окисления углеводов на основании ДК будет неточным, если энергия освобождается за счет этого процесса.
Несмотря на определенные недостатки метода непрямой калориметрии, он остается лучшим способом определения расхода энергии в покое и при субмаксимальных нагрузках.
ИЗОТОПНЫЕ ИЗМЕРЕНИЯ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА
В прошлом определение общего расхода энергии за день осуществлялось на основании тщательной регистрации потребления пищи в течение нескольких дней, а также измерений изменений в составе тела за этот период. Этот метод, несмотря на широкое применение, был ограничен способностью человека вести точную регистрацию и точно определять расход энергии мышечной деятельности.
Применение изотопов расширило наши возможности изучения обмена энергии. Изотопы представляют собой элементы с нетипичной атомной массой. Они бывают радиоактивными (радиоизотопы) или нерадиоактивными (устойчивые изотопы). Например, углерод-12 (^С) имеет молекулярную массу 12 и является наиболее естественной формой углерода. Он нерадиоактивен. Существует также углерод-14 ('''С), у которого на два нейтрона больше, чем у углерода-12, поэтому его атомная масса 14. Углерод-14 образуется в лабораторных условиях и является радиоактивным.
Углерод-13 (^С) составляет 1 % углерода в природе и очень часто используется для изучения обмена энергии. Он нерадиоактивен, поэтому за ним труднее следить в организме, чем за углеродом-14. Хотя за радиоактивными изотопами легко следить в организме, они отрицательно влияют на ткани тела и поэтому их редко используют в исследованиях с участием людей.
Углерод-13 и другие изотопы, например, во-
дород-2 (дейтерий, или 1Н) используются как меченые атомы, т.е. за их перемещениями в организме человека можно наблюдать. Этот метод предполагает введение изотопов человеку и последующее слежение за их распределением и перемещением.
Хотя данный метод описан более 30 лет назад, исследования с использованием меченой воды стали проводиться лишь совсем недавно для контроля расхода энергии в обычных условиях. Испытуемый выпивает известное количество воды, меченой двумя изотопами ('Н^Ю). Дейтерий (^Н) диффундирует в жидкости организма, а кислород-18 ('Ю) — в жидкости, а также в запасы двууглекислой соли (где содержится большая часть диоксида углерода, образованного вследствие метаболизма). Интенсивность выделения двух изотопов из организма можно определить, анализируя их присутствие в моче, слюне или пробах крови. На основании полученных данных можно определить количество образуемого СО^ и затем, используя калориметрические уравнения, перевести полученный показатель в величину расходуемой энергии.
Поскольку обмен изотопов происходит относительно медленно, измерения обмена энергии следует проводить в течение нескольких недель. Следовательно, этот метод не совсем подходит для определения срочного метаболизма. Вместе с тем, благодаря высокой (свыше 98 %) точности и небольшой степени риска его можно считать весьма приемлемым для определения затрат энергии, происходящих изо дня в день. Специалисты в области питания утверждают, что данный метод представляет собой наиболее значительное техническое открытие прошлого столетия в области обмена энергии.
В ОБЗОРЕ...
1. Метод прямой калориметрии основан на использовании калориметра для непосредственного измерения количества тепла, образуемого телом.
2. Метод непрямой калориметрии предполагает измерение потребления 0^ и выделения СОд, вычисление ДК (соотношение показателей этих газов) и его сопоставление со стандартными показателями для определения вида окисленного субстрата и последующего определения расхода энергии на литр поглощенного кислорода.
3. Обычные значения ДК — 0,78 — 0,80.
4. Для определения интенсивности метаболизма могут использоваться изотопы. Их вводят в организм и следят за их перемещениями. Скорость их выделения позволяет определить образование СО^ и расход энергии.
ОЦЕНКА АНАЭРОБНОГО УСИЛИЯ
Мы рассмотрели способы определения аэробного метаболизма, однако они не учитывают анаэробные процессы. Как можно оценить взаимодействие аэробных (окислительных) и анаэробных процессов? Наиболее распространенными методами оценки анаэробного усилия являются методы, включающие изучение избыточного потребления кислорода после нагрузки либо лак-татного порога. Рассмотрим оба метода.
Дата: 2018-12-28, просмотров: 403.