ЭНЕРГИЯ ДЛЯ КЛЕТОЧНОЙ ДЕЯТЕЛЬНОСТИ
Источником всех видов энергии является энергия Солнца. Химические реакции, происходящие в растениях (фотосинтез), превращают энергию Солнца в химическую энергию. Употребляя в пищу овощи и фрукты, а также мясо животных, которые питаются растениями, мы получаем энергию. Энергия содержится в пищевых продуктах в виде углеводов, жиров и белков. Эти основные компоненты пищевых продуктов расщепляются в клетках нашего организма, освобождая энергию.
Поскольку энергия постепенно превращается в тепло, ее количество, высвобождаемое в результате биологических реакций, определяется по количеству образованного тепла. В биологических
85
системах энергия измеряется в килокалориях (ккал). Согласно определению, 1 ккал равна количеству тепловой энергии, необходимой для повышения температуры 1 кг воды с 1 до 15 °С. Например, при сжигании спички выделяется примерно 0,5 ккал, тогда как при полном сгорании 1 г углеводов выделяется 4,0 ккал.
Некоторое количество свободной энергии в клетках используется для развития и "ремонта" нашего организма. Такие процессы, как мы уже знаем, направлены на развитие мышечной массы под влияние тренировочных нагрузок и восстановление мышц после физических нагрузок или травм. Энергия также необходима для активного транспорта множества веществ, таких, как глюкоза и Са24' через клеточные мембраны. Активный транспорт необходим для функционирования клеток и поддержания гомеостаза. Определенное количество энергии используют миофибриллы для обеспечения скольжения филаментов актина и миозина, в результате которого производятся мышечное сокращение и сила, что мы уже рассматривали в главе 2.
ИСТОЧНИКИ ЭНЕРГИИ
Продукты питания состоят, в основном, из углерода, водорода, кислорода, а при наличии белков — из азота. Молекулярные связи в пищевых продуктах относительно слабые, и в случае расщепления освобождается небольшое количество энергии. Следовательно, продукты питания не используются непосредственно для потребностей клетки. Энергия молекулярных связей продуктов питания химически освобождается в клетках организма и хранится в виде высокоэнергетического соединения — аденозинтрифосфата (АТФ).
'у Образование АТФ позволяет клеткам со-т хранять энергию в этом высокоэнергети
Ческом соединении
В покое энергия, необходимая нашему организму, обеспечивается за счет расщепления практически одинакового количества углеводов и жиров. Белки представляют собой "строительные блоки" и обычно обеспечивают функционирование клеток небольшим количеством энергии. При увеличении мышечного усилия в качестве источника энергии больше используются углеводы. При максимальной кратковременной нагрузке АТФ почти исключительно образуется за счет углеводов.
Углеводы
Зависимость мышц от углеводов во время физической нагрузки связана с их наличием, а также способностью мышечной системы их расщеплять.
Содержание углеводов в печени и скелетных мышцах ограничено; их достаточно для образования не более 2 000 ккал энергии. Это количество расходуется на то, чтобы пробежать 32 км (20 миль). Запасы жиров достаточны для образования более 70 000 ккал энергии
Углеводы в конечном итоге превращаются в глюкозу — моносахарид, который транспортируется кровью ко всем тканям организма. В состоянии покоя поглощенные углеводы попадают в мышцы и печень, а затем преобразуются в более сложную молекулу сахара — гликоген. Гликоген находится в цитоплазме до тех пор, пока клетки не используют его для образования АТФ. Гликоген, содержащийся в печени, может снова превращаться в глюкозу, она транспортируется кровью к активным тканям, где и происходит ее метаболизм (расщепление).
Содержание гликогена в печени и мышцах ограничено, и его запасы могут истощиться, если в рационе питания нет достаточного количества углеводов. Таким образом, восполнение запаса углеводов во многом зависит от пищевых источников крахмалов и Сахаров. Без достаточного потребления углеводов мышцы и печень лишаются своего основного источника энергии.
Жиры
Жиры и белки также используются в качестве источников энергии. В организме содержится значительно больше жиров, чем углеводов. Как видно из табл. 5.1, жировой резерв энергии в организме значительно превышает углеводный. Однако жиры менее доступны клеточному метаболизму, поскольку прежде всего должна быть расщеплена сложная форма — триглицерид — на основные компоненты: глицерин и свободные жирные кислоты. Только свободные жирные кислоты используются для образования АТФ.
Таблица 5.1. Запасы "топлива" и энергии в организме
| Источник энергии | г | ккал |
| Углеводы гликоген печени мышечный гликоген глюкоза в жидкостях организма Всего Жиры подкожный внутримышечный Всего
| 110 250 15 |
0,451 1,025 0,062
1,538 70,980 1,465
|
| 375 7,800 0,161 | ||
| 7,961 72,445
| ||
| Примечание. Оценка произведена на основании средней массы тела 65 кг с содержанием жира 12 %.
| ||
86
Как видно из рис. 5.1, из одного и того же количества жиров и углеводов образуется абсолютно разное количество энергии: соответственно 9 и 4 ккалт'. В любом случае интенсивность освобождения энергии из этих соединений слишком небольшая, чтобы удовлетворить потребности организма в энергии во время интенсивной мышечной деятельности.
Белки
Процесс превращения белков или жиров в глюкозу называется глюконеогенезом. В результате серии реакций белок может превратиться в жирные кислоты. Это — липогенез.
Белки обеспечивают 5 — 10 % энергии, необходимой для выполнения продолжительного физического упражнения. Для образования энергии используются лишь основные единицы белка-аминокислоты.
ИНТЕНСИВНОСТЬ ВЫСВОБОЖДЕНИЯ ЭНЕРГИИ
Чтобы быть полезной, энергия должна высвобождаться из химических соединений с контролируемой интенсивностью. Частично эта интенсивность определяется выбором источника энергии. Если используется большое количество энергии из одного источника, клетки рассчитывают главным образом именно на этот источник. Такое влияние наличия энергии называется эффектом массового воздействия.
Специальные ферменты обеспечивают более четкий контроль интенсивности высвобождения свободной энергии. Многие из них облегчают расщепление (катаболизм) химических соединений (рис. 5.2). Хотя названия ферментов довольно сложны, все они заканчиваются суффиксом -аза. Например, фермент, воздействующий на АТФ, называется аденозинтрифосфатаза (АТФаза).
Разобравшись с источниками энергии, рассмотрим, как сохраняется энергия. В следующем параграфе мы изучим, как образуется содержащее энергию соединение — АТФ.
Жиры
(свободные жирные кислоты)
| Углеводы |
 1 г С6Н12О6 1 г С„НзА
1 г С6Н12О6 1 г С„НзА
 4 ккал энергии 9 ккал энергии
4 ккал энергии 9 ккал энергии
В ОБЗОРЕ...
1. Около 60 — 70 % энергии в организме человека превращается в тепло. Остальное количество используется для выполнения механической работы и осуществления клеточной деятельности.
2. Мы получаем энергию из пищевых продуктов — углеводов, жиров и белков.
3. Получаемая из продуктов питания энергия содержится в высокоэнергетическом соединении — АТФ.
4. Углеводы обеспечивают около 4 ккал энергии на 1 г, тогда как жиры —около 9 ккалт"1, однако энергия, получаемая из углеводного источника, более доступна. Белки также обеспечивают организм энергией.
Молекула АБ
| Фермент |
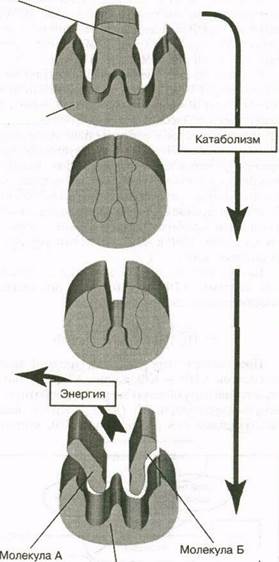
Рис. 5.1. Образование энергии из 1 г углеводов и ] г жиров
Фермент
Рис. 5.2. Действие ферментов в катаболизме (расщеплении) соединений
87
| Рис. 5.3 Структурный состав молекулы АТФ с энергобогатыми фосфатными соединениями (а) и высвобождение энергии (б) |
а ^АТФ^ = | Аденозин |— Энергия —(Р.)— Энергия —(Р.)—— Энергия —(Р.)
. АТФаза +
б | Аденозин~|-{р)—(Р}—(Р) ^—-^| Аденозин | - (Р)—(Р) + (р) + Энергия
АДФ
АТФ
БИОЭНЕРГЕТИКА: ОБРАЗОВАНИЕ АТФ
Молекула АТФ (рис. 5.3,а) состоит из аденози-на (молекулы аденина, соединенной с молекулой рибозы), соединенного с тремя группами неорганического фосфата (Р ). При воздействии фермента АТФазы последняя фосфатная группа отщепляется от молекулы АТФ, быстро высвобождая большое количество энергии (7,6 ккал-моль'' АТФ). В результате АТФ расщепляется на АДФ (аденозин-дифосфат) и фосфор (рис. 5.3,6). Однако где же находилась эта энергия в самом начале?
Процесс накопления энергии в результате образования АТФ из других химических источников называется фосфорилированием. Вследствие различных химических реакций фосфатная группа присоединяется к относительно низкоэнергетическому соединению — аденозиндифосфату, преобразуя его в аденозинтрифосфат. Когда эти реакции осуществляются без наличия кислорода, процесс называется анаэробным метаболизмом. Если же в реакции участвует кислород, процесс называется аэробным метаболизмом, а аэробное превращение АДФ в АТФ —окислительным фосфорилированием.
Клетки образуют АТФ с помощью трех систем: системы АТФ — КФ, гликолитической и окислительной систем.
СИСТЕМА АТФ - КФ
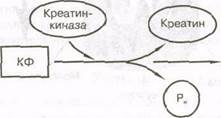
Простейшей энергетической системой является система АТФ — КФ. Кроме АТФ, клетки содержат еще одну богатую энергией фосфатную молекулу —креатинфосфат (КФ). Энергия, высвобождающаяся при расщеплении КФ, в отличие
от энергии, высвобождающейся при расщеплении АТФ, не используется непосредственно для выполнения работы на клеточном уровне. Она используется для ресинтеза АТФ, чтобы обеспечить его относительно постоянное образование.
Высвобождению энергии при расщеплении КФ содействует фермент креатинкиназа, который действует на КФ с целью отделения Р^ от креатина. Освобожденная энергия может быть использована для присоединения Р^ к молекуле АДФ и образования АТФ (рис. 5.4). При использовании этой системы (энергия высвобождается из АТФ в результате отщепления фосфатной группы) клетки могут предотвратить истощение запасов АТФ, расщепляя КФ, и тем самым обеспечивая энергию для образования большего количества АТФ.
Это быстрый процесс, который может осуществляться без помощи каких-либо специальных структур клетки. Он может протекать и с участием кислорода, однако для его осуществления кислород не нужен, поэтому систему АТФ — КФ называют анаэробной.
В первые секунды интенсивной мышечной деятельности количество АТФ поддерживается на относительно постоянном уровне, тогда как уровень КФ неуклонно снижается, поскольку он используется для пополнения запасов АТФ (рис. 5.5). В состоянии изнеможения уровни АТФ и КФ довольно низки и не могут обеспечить энергию для последующих сокращений и расслаблений мышц.
Таким образом, поддержание уровня АТФ за счет энергии, высвобождающейся при расщеплении КФ, ограничено. Запасы АТФ и КФ достаточны для удовлетворения энергетических потребностей мышц лишь в течение 3 — 15с спринтер-
|
|
| Энергия |
|
|
| ( аТФ) |
Рис. 5.4
Поддержание уровня АТФ за счет энергии,
содержащейся вКФ
88
| 100 |
| 80 |
| 60 |
| 40 |
| 20 |
2 4 6 8 10 12 14
Время,с
Рис. 5.5. Изменения в мышечных АТФ и КФ в первые секунды максимального мышечного усилия
ского бега. После этого мышцам приходится рассчитывать на другие процессы образования АТФ:
гликолитический и окислительный.
ГЛИКОЛИТИЧЕСКАЯ СИСТЕМА
Другой источник получения АТФ предусматривает высвобождение энергии в результате расщепления (лизиса) глюкозы. Это — гликолитичес-кая система, включающая процесс гликолиза, т.е. расщепление глюкозы с помощью специальных гликолитических ферментов (рис. 5.6).
Глюкоза составляет около 99 % всех Сахаров, циркулирующих в крови. Она поступает в кровь в результате усвоения углеводов и расщепления гликогена печени. Гликоген синтезируется из глюкозы вследствие процесса, называемого гликогенезом. Гликоген содержится в печени или мышцах до тех пор, пока не потребуется. Когда возникает потребность в гликогене, он расщепляется в результате процесса гликогенолиза на глюкозо-1 -фосфат.
Прежде чем глюкоза или гликоген могут быть использованы для образования энергии, они должны трансформироваться в соединение, которое называется глюкозо-6-фосфат. Для превращения молекулы глюкозы необходима одна молекула АТФ. При расщеплении гликогена глюкозо-6-фосфат образуется из глюкозо-1 -фосфата без затраты энергии. Гликолиз начинается, как только образуется глюкозо-6-фосфат.
Заканчивается гликолиз образованием пиро-виноградной кислоты. Для этого процесса не нужен кислород, однако использование кислорода определяет "судьбу" пировиноградной кислоты, образованной вследствие гликолиза. Когда мы говорим о гликолитической системе, мы имеем в виду, что процесс гликолиза протекает без участия кислорода. В этом случае пировиноградная кислота превращается в молочную кислоту.
Гликолиз, являясь более сложным процессом, чем система АТФ — КФ, обеспечивает расщеп
ление гликогена на молочную кислоту благодаря 12 ферментным реакциям. Все эти ферменты находятся в цитоплазме клеток. В результате гликолиза образуется 3 моля АТФ на каждый моль расщепленного гликогена. Если вместо гликогена используется глюкоза, образуется всего 2 моля АТФ, поскольку один моль расходуется на превращение глюкозы в глюкозо-6-фосфат.
Эта энергетическая система не обеспечивает образование большого количества АТФ. Несмотря на это, сочетанные действия гликолитической системы и системы АТФ — КФ обеспечивают производство силы мышцами даже при ограниченном поступлении кислорода. Эти две системы доминируют в первые минуты выполнения упражнений высокой интенсивности.
Другим значительным недостатком анаэробного гликолиза является то, что он вызывает накопление молочной кислоты в мышцах и жидкостях организма. В спринтерских дисциплинах продолжительностью 1 — 2 мин потребности гликолитической системы высоки, и уровни содержания молочной кислоты могут увеличиться с 1 (показатель в состоянии покоя) до более чем 25 ммоль-кг'. Такое подкисление мышечных волокон тормозит дальнейшее расщепление гликогена, поскольку нарушает функцию гликолитических ферментов. Кроме того, кислота снижает способность волокон связывать кальций и это может препятствовать сокращению мышц.
Интенсивность энергозатрат мышечного во-
Глюкоза Гликоген

Глюкозо-6-фосфат
 Гликолитические ферменты
Гликолитические ферменты
 АТФ
АТФ
 Пировиноградная кислота
Пировиноградная кислота
 Молочная кислота
Молочная кислота
Рис. 5.6. Процесс гликолиза
99
Молочная кислота и лактат — не одно и то же соединение. Молочная кислота имеет формулу СдНдОд. Лактат представляет собой любую соль молочной кислоты. Когда молочная кислота теряет Н^ оставшееся соединение, соединяясь с Nа+ или К^ образует соль. В результате анаэробного гли-колиза образуется молочная кислота, которая очень быстро разлагается и образует соль — лактат. Из-за этого очень часто одно понятие используют вместо другого
локна во время нагрузки может быть в 200 раз выше, чем в состоянии покоя. Гликолитическая система и система АТФ — КФ не в состоянии обеспечить необходимое количество энергии. Без другой энергетической системы наша способность выполнять мышечную деятельность была бы ограничена всего несколькими минутами. Рассмотрим третью энергетическую систему.
ОКИСЛИТЕЛЬНАЯ СИСТЕМА
Последней системой образования энергии клеткой является окислительная система, наиболее сложная из трех энергетических систем. При ее рассмотрении мы опустим обременительные подробности. Процесс, в результате которого организм для производства энергии диссимилирует соединения, богатые энергией, с помощью кислорода называется клеточным дыханием. Это аэробный процесс, поскольку в нем участвует кислород. АТФ образуется в специальных клеточных органеллах — митохондриях. В мышцах они примыкают к миофибриллам, а также разбросаны по саркоплазме.
Мышцам необходимо постоянное обеспечение энергией для производства силы во время продолжительной мышечной деятельности. В отличие от анаэробного образования АТФ окислительная система производит значительное количество энергии, поэтому аэробный метаболизм — основной метод образования энергии во время мышечной деятельности, требующей проявления выносливости. Это предъявляет повышенные требования к системе транспорта кислорода к активным мышцам.
В ОБЗОРЕ...
1. Три энергетические системы обеспечивают образование АТФ:
• система АТФ — КФ;
• гликолитическая система;
• окислительная система.
2. В системе АТФ — КФ Р^ отделяется от кре-атинфосфата под действием креатинкиназы. Р^ затем может соединиться с АДФ, образуя АТФ. Это анаэробная система, главная функция которой — поддержание уровня АТФ. Величина высвобождения энергии составляет 1 моль АТФ на 1 моль КФ.
3. Гликолитическая система включает процесс гликолиза, посредством которого глюкоза или гликоген расщепляется на пировиноградную кислоту с помощью гликолитических ферментов. Если в процессе не участвует кислород, пирови-ноградная кислота превращается в молочную кислоту. Из 1 моля глюкозы образуется 2 моля АТФ, тогда как из 1 моля гликогена — 3 моля АТФ.
4. Гликолитическая система и система АТФ — КФ — основные источники энергии в первые минуты выполнения физического упражнения высокой интенсивности.
Окисление углеводов
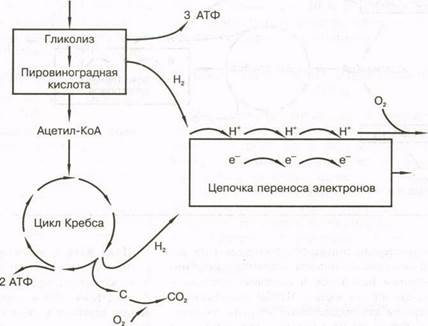
Окислительное образование АТФ (рис. 5.7) включает три процесса:
1)гликолиз;
2) цикл Кребса;
3) цепочку переноса электронов.
Гликолиз при обмене углеводов играет важную роль как в анаэробном, так и аэробном образовании АТФ. Причем он протекает одинаково, независимо от того, участвует ли в процессе кислород. Участие кислорода определяет лишь "судьбу" конечного продукта — пировиноградной кислоты. Вспомним, что при анаэробном глико-лизе образуется молочная кислота и всего 3 моля АТФ на 1 моль гликогена. При участии кислорода пировиноградная кислота превращается в соединение, которое называется ацетил-кофермент А (ацетил-КоА).
Цикл Кребса. После образования ацетил-КоА попадает в цикл Кребса (цикл лимонной кислоты) — сложную последовательность химических реакций, которые позволяют завершить окисление ацетил-КоА. В конце цикла Кребса образуется 2 моля АТФ, а вещество (соединение, на которое воздействуют ферменты, в данном случае — первоначальный углевод) расщепляется и соединяясь с кислородом, образует диоксид углерода (СО-), который легко диффундирует из клеток, транспортируется кровью в легкие и выделяется во внешнюю среду (пост реакция декарбоксили-рования К - Н + СО,).
Цепочка переноса электронов. Во время гликолиза, когда глюкоза превращается в пировиноградную кислоту, выделяется водород. Значительно большее количество водорода выделяется во время цикла Кребса. Если он остается в системе,
90
| Рис. 5.7 Окисление углеводов |
| СНО (гликоген) |
 |
Глюкозо-6-фосфат
|
внутренняя часть клеток становится слишком кислой. Что происходит с этим водородом?
Таблица 5.2 Образование энергии вследствие окисления гликогена печени
| АТФ, образуемый из 1 моля
| ||
| гликогена печени
| ||
| Этап процесса | окислительным | |
| непосредственно | фосфорилированием* | |
| Гликолиз 3
| 6 | |
| (превращение
| ||
| глюкозы в пиро-
| ||
| виноградную
| ||
| кислоту)
| ||
| Превращение 0
| 6 | |
| пировиноградной
| ||
| кислоты
| ||
| в ацетил-КоА
| ||
| Цикл Кребса 2
| 22 | |
| 5
| 34 | |
| Итого 39
| ||
| * Относится к АТФ, образованному при переносе Н* И электронов в цепочку переноса электронов.
| ||
Цикл Кребса связан с серией реакций, которые называются цепочкой переноса электронов. Водород, выделяющийся во время гликолиза и в цикле Кребса, соединяется с двумя коферментами — ни-котинамид-аденин-динуклеотидом и флавин-аде-нин-динуклеотидом, которые переносят атомы во
дорода в цепочку переноса электронов, где они расщепляются на протоны и электроны. В конце цепочки Н"1" соединяется с кислородом, образуя воду и тем самым предотвращая подкисление.
Электроны, отделившиеся от водорода, принимают участие в серии реакций и в конечном итоге обеспечивают энергию для фосфорилиро-вания АДФ, а следовательно, образования АТФ. Поскольку этот процесс проходит с участием кислорода, он называется окислительным фосфорилированием.
Образование энергии за счет углеводов. Окислительная система образования энергии обеспечивает получение 39 молекул АТФ из одной молекулы глюкозы. Если процесс начинается с глюкозы, образуется 38 молекул АТФ (вспомним, что одна молекула АТФ используется до начала гликолиза для образования глюкозо-фосфата). В табл. 5.2 показано количество образующейся энергии.
Окисление жиров
Как уже отмечалось, жиры также вносят свой вклад в энергетические потребности мышц. Запасы гликогена в мышцах и печени могут обеспечить всего 1 200 — 2 000 ккал энергии, в то время как жиры, содержащиеся внутри мышечных волокон и в жировых клетках, — около 70 000 — 75 000 ккал.
Хотя жирами называются многие химические соединения, такие, как триглицериды, фосфоли-
91
|
|

пиды и холестерин, только триглицериды используются в качестве основного источника энергии. Триглицериды находятся в жировых клетках и волокнах скелетных мышц. Чтобы использовать триглицериды для образования энергии, необходимо расщепить их на основные составляющие:
одну молекулу глицерина и три молекулы свободных жирных кислот. Этот процесс называется липолизом и осуществляется ферментами — липазами. Поскольку свободные жирные кислоты — основной источник энергии, мы уделим им главное внимание.
Высвободившись из триглицерида (жира), свободные жирные кислоты могут попасть в кровь, которая транспортирует их по всему организму, и в результате диффузии проникнуть в мышечные волокна. Интенсивность поступления свободных жирных кислот в мышечные волокна зависит от градиента концентрации. Повышение концентрации свободных жирных кислот в крови выталкивает их в мышечные волокна.
B-Окисление. Несмотря на значительные структурные различия между разными свободными жирными кислотами, их метаболизм почти одинаков, как следует из рис. 5.8. До того, как они попадут в мышечные волокна, свободные жирные кислоты активируются энергией АТФ с помощью ферментов. Таким образом их подготавливают к катаболизму (расщеплению) в митохон-дриях. Этот ферментный катаболизм жиров митохондриями называется бета-окислением.
В этом процессе углеродная цепочка свободной жирной кислоты делится на двухуглеродные остатки уксусной кислоты. Например, если первоначально свободная жирная кислота имела 16-углеродную цепочку, то при (3-окислении образуется 8 молекул уксусной кислоты. Вся уксусная кислота затем превращается в ацетил-КоА.
Таблица 5.3. Образование энергии вследствие окисления пальмитиновой кислоты (С,дЦцО;)
АТФ, образуемый из 1 молекулы пальмитиновой кислоты
Этап процесса
окислительным фосфорилированием
| непосредственно |
Активация жирной
| -2 |
35
88 121
кислоты
B-Окисление
Цикл Кребса 8 / 8
Итого
129
МЕТАБОЛИЗМ БЕЛКОВ
Как уже отмечалось, углеводы и жиры — предпочтительные источники энергии нашего организма. Однако используются и белки или, скорее, аминокислоты, из которых они состоят. Некоторые аминокислоты могут превращаться в глюкозу (посредством глюконеогенеза). Другие могут превращаться в различные промежуточные продукты окислительного метаболизма (такие, как пируват или ацетил-КоА), чтобы принять участие в окислительном процессе.
Количество энергии, образуемой белками, довольно трудно определить в отличие от энергии, образуемой углеводами или жирами, поскольку белки также содержат азот. При катаболизме аминокислот некоторое количество азота используется для образования новых аминокислот, остальное количество азота превращается в мочевину и выделяется главным образом с мочой. Этот процесс требует использования АТФ и, следовательно, приводит к затратам некоторого количества энергии.
При расщеплении белков посредством сжигания в лабораторных условиях образуется 5,65 ккалт"' энергии. При метаболизме белка в организме вследствие затрат энергии на процесс превращения азота в мочевину высвобождается всего 5,20 ккалт"' энергии, т.е. на 8 % меньше.
Чтобы точно определить интенсивность обмена белков, необходимо установить, какое коли
чество азота выводится из организма. Для этого необходимо собирать мочу в течение 12— 24 ч. Поскольку здоровый организм использует небольшое количество белков в состоянии покоя и при выполнении физической нагрузки (как правило, намного меньше 5 — 10 % всех затрат энергии), при оценке затрат энергии метаболизм белков просто не принимают во внимание.
ОКИСЛИТЕЛЬНЫЕ СПОСОБНОСТИ
МЫШЦ
Мы выяснили, что процессы окислительного метаболизма обеспечивают максимальное образование энергии. Было бы идеальным, если бы эти процессы всегда осуществлялись с максимальной эффективностью. Однако, как и все физиологические системы, они имеют свои ограничения. Окислительная способность (?о мышцы — показатель ее максимальной способности использовать кислород. В следующем параграфе мы рассмотрим факторы, ограничивающие эту способность мышц.
Активность ферментов
Способность мышечных волокон окислять углеводы и жиры довольно трудно определить. Во многих исследованиях наблюдали тесную взаимосвязь между способностью мышцы выполнять аэробное упражнение в течение длительного времени и активностью ее окислительных ферментов. Поскольку для окисления требуется много ферментов, их активность в мышечных волокнах — достаточно надежный показатель окислительного потенциала.
Нет смысла измерять все ферменты, поэтому для иллюстрации аэробной способности волокон выбирают некоторые из них. Чаше всего используют такие ферменты, как сукцинатдегидрогена-за и цитратсинтаза, ферменты митохондрий, участвующие в цикле Кребса. На рис. 5.9 проиллюстрирована взаимосвязь между активностью сукцинатдегидрогеназы в латеральной широкой мышце и ее окислительной способностью. Окислительная активность ферментов в мышцах спортсменов, занимающихся видами спорта, требующими проявления выносливости, в два — четыре раза выше, чем в мышцах нетренированных мужчин и женщин [2, 3, 5].
Рис. 5.12
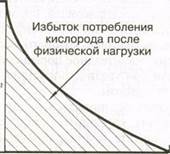
Потребность в кислороде во время выполнения физической нагрузки и во время восстановления. Показаны дефицит кислорода и избыток потребления кислорода после физической нагрузки
| П О Т Р Е Б Н Ость О2 |
| Потребность в О; |
| Дефицит О; |
| Потребление О; в покое |
|
|
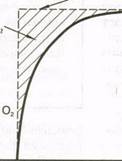
| Устойчивое потребление О; |
| Начало нагрузки |
| Конец нагрузки Время |
|
|
| Конец восстановления |
 |
видных компонента: исходный быстрый и вторичный медленный. Согласно классической теории, быстрый компонент кривой отражает количество кислорода, необходимого для восстановления АТФ и КФ, используемых во время физической нагрузки, особенно в самом начале. Без достаточного количества кислорода макро-энергетические фосфатные связи в этих соединениях нарушаются, вследствие чего не поставляется необходимое количество энергии. Во время восстановления эти связи должны быть возобновлены благодаря окислительным процессам, чтобы восполнить запасы энергии или устранить долг. Считалось, что медленный компонент кривой — результат выведения аккумулированного лактата из тканей путем либо превращения в гликоген, либо окисления в СО^ и НдО, что обеспечивает энергию, необходимую для восстановления запасов гликогена.
В рамках этой теории полагали, что оба компонента кривой отражают анаэробную деятельность во время физической нагрузки. Существовало мнение, что на основании потребления кислорода после выполнения упражнения можно определить величину анаэробной производительности.
Недавние исследования, однако, показали, что классическое объяснение избыточного потребления кислорода слишком упрощено. Например, вначале выполнения упражнения некоторое количество кислорода может быть взято из запасов (гемоглобина и миоглобина). Во время восстановления это количество кислорода должно быть восполнено. Кроме того, после выполнения упражнения дыхание некоторое время остается учащенным. Частично это обусловлено попыткой устранить накопившийся в тканях СО- как продукт метаболизма. Температура тела также повышена, в результате чего поддерживается высокая интенсивность метаболизма и дыхания, что требует больше кислорода. Точно так же влияют повышенные уровни адреналина и норадреналина.
Таким образом, классическая теория не учитывает все факторы, от которых зависит кислородный долг. Необходимо более тщательное изу
чение физиологических механизмов, обусловливающих избыток потребления кислорода после физической нагрузки.
Порог лактата
Многие специалисты считают порог лактата надежным показателем потенциальных возможностей спортсмена выполнять физические нагрузки, требующие проявления выносливости. Порог лактата определяют как момент начала аккумуляции лактата в крови во время физической нагрузки увеличивающейся интенсивности сверх уровней, характерных для состояния покоя. Если интенсивность мышечной деятельности небольшая или средняя, уровень лактата лишь немного превышает показатель в состоянии покоя. Увеличение интенсивности приводит к более быстрой аккумуляции лактата. При невысокой скорости плавания (рис. 5.13) уровни лактата равны или близки к уровням, характерным для состояния покоя. При увеличении скорости плавания более 1,4 м-с'' уровни лактата крови быстро повышаются. Эта точка разрыва непрерывности на кривой соответствует порогу лактата.
Согласно определению, порог лактата должен
7* 99
отражать взаимодействие между аэробной и анаэробной энергетическими системами. По мнению некоторых исследователей, порог лактата отражает значительный сдвиг в сторону анаэробного гли-колиза, вследствие которого образуется лактат. Поэтому значительное повышение уровня лактата крови при увеличении усилия называют анаэробным порогом.
Следует остановиться на значительных противоречиях, касающихся взаимосвязи между порогом лактата и анаэробным метаболизмом в мышце. Мышцы, безусловно, образуют лактат еще до того, как достигается порог лактата, однако его выводят другие ткани. Кроме того, точка разрыва непрерывности не всегда очевидна. Поэтому специалисты довольно часто устанавливают произвольный показатель 2,0 или 4,0 ммоль лактата на литр потребляемого кислорода, отражающий момент начала аккумуляции лактата крови. Это —эталонный момент начала аккумуляции лактата крови или отправной момент работы.
У нетренированных людей порог лактата обычно наблюдается при 50— 60 % МПК. У сильнейших спортсменов, занимающихся видами спорта, требующими проявления выносливости, порог лактата достигается при 70- 80% МП К
повышенную потребность в кислороде после выполнения физической работы.
3. Порог лактата — это момент, когда происходит быстрая аккумуляция лактата в крови, во время выполнения физической нагрузки, превышающая уровни лактата в покое'.
4. Как правило, люди, имеющие более высокие показатели порога лактата или начала аккумуляции лактата крови, выраженные в процентном отношении к МПК, способны показать более высокие результаты в видах спорта, требующих проявления выносливости.
Порог лактата, выраженный в процентном отношении к МПК — один из лучших показателей тренированности спортсмена в таких циклических видах спорта, как бег на длинные дистанции и езда на велосипеде
| Т |
Порог лактата обычно выражают в процентах МПК, при котором он достигается. Способность выполнять работу с высокой интенсивностью без накопления лактата имеет большое значение, поскольку образование лактата способствует утомлению. Следовательно, порог лактата при 80 % МПК свидетельствует о лучшей толерантности к физическим нагрузкам по сравнению с лактат-ным порогом при 60 % МПК. Из двух людей с одинаковыми МПК более высокий уровень физической деятельности, требующей проявления выносливости, будет у того, у которого выше порог лактата.
В ОБЗОРЕ...
1. Избыточное потребление кислорода после физической нагрузки представляет собой повышенное по сравнению с состоянием покоя потребление его, которое продолжается некоторое время после завершения физической нагрузки.
2. Традиционно считалось, что избыточное потребление кислорода отражает анаэробное усилие во время выполнения физической нагрузки, однако это слишком упрощенное представление. Сочетание целого ряда факторов обусловливает
100
Этот показатель соответствует средним затратам энергии в состоянии покоя у человека массой 70 кг (154 фунта). Естественно, он не включает количество энергии, необходимой для выполнения обычной повседневной деятельности.
Стандартная мера затрат энергии в покое — скорость основного обмена, т.е. скорость энергозатрат у человека в состоянии покоя в положении лежа на спине, которая измеряется сразу после сна в течение не менее 8 ч и после голодания в течение не менее 12 ч. Это показатель отражает минимальное количество энергии, необходимой для осуществления физиологических функций организма.
Скорость основного обмена непосредственно связана с чистой массой тела и обычно выражается в килокалориях на килограмм чистой массы тела в минуту. Чем выше чистая масса тела, тем больше килокалорий расходуется за день. Вспомним, что у женщин масса жира больше, чем у мужчин. Поэтому у них, как правило, показатели скорости основного обмена ниже, чем у мужчин одинаковой с ними массы.
Такое же большое значение имеет и площадь поверхности тела. Чем больше площадь поверхности тела, тем больше отдача тепла телом и выше скорость основного обмена, поскольку для поддержания температуры тела требуется больше энергии. Поэтому скорость основного обмена часто выражают в килокалориях на 1 м2 площади поверхности тела в час. Поскольку мы обсуждаем расход энергии за день, используем более подходящую единицу —килокалории в день.
На скорость основного обмена влияет множество других факторов. Среди них:
• возраст: скорость основного обмена с возрастом постепенно снижается;
• температура тела: скорость основного обмена увеличивается при повышении температуры;
• стресс: стресс повышает активность симпатической нервной системы, что, в свою очередь, увеличивает скорость основного обмена;
• гормоны: тироксин щитовидной железы и адреналин мозгового вещества надпочечников повышают скорость основного обмена.
В настоящее время вместо скорости основного обмена большинство специалистов используют термин "интенсивность обмена веществ в покое", поскольку большинство измерений осуществляется таким же образом, как и при определении скорости основного обмена. Показатели скорости основного обмена колеблются от 1 200 до 2 400 ккал-день-'. Общая средняя интенсивность метаболизма при обычной повседневной деятельности колеблется от 1 800 до 3 000 ккал.
Расход энергии у крупных спортсменов, ежедневно проводящих интенсивные тренировки, может превышать 10 000 ккал-день~Ч
МАКСИМАЛЬНАЯ СПОСОБНОСТЬ ВЫПОЛНЯТЬ ФИЗИЧЕСКИЕ НАГРУЗКИ
При переходе от состояния покоя к выполнению физической нагрузки потребность в энергии возрастает. Метаболизм увеличивается прямо пропорционально увеличению интенсивности работы. Сталкиваясь с возрастающими потребностями в энергии, наш организм постепенно достигает предела потребления кислорода. В этот момент, как следует из рис. 5.14, потребление кислорода (То,) достигает пика и остается постоянным или даже слегка снижается, несмотря на все возрастающую интенсивность работы. Этот пиковый показатель представляет собой аэробную способность, максимальное потребление кислорода или МПК. По мнению ряда ученых, МПК — лучший показатель уровня кардиореспираторной выносливости и аэробной подготовленности.
Хотя, по мнению некоторых специалистов в области спорта, МПК — хороший прогностический фактор успеха в видах спорта, требующих проявления выносливости, тем не менее, на основании только этого показателя, определенного в лабораторных условиях, нельзя предопределить победителя марафонского забега [4]. Точно так же тест бега на длинную дистанцию может лишь приблизительно предсказать индивидуальный показатель МПК. Это свидетельствует о том, что хороший спортивный результат зависит не только от высокого МПК [I].
Исследования показывают, что МПК увеличивается вследствие физических тренировок толь-
ко в течение 8—12 недель, затем наблюдается плато, несмотря на дальнейшее увеличение интенсивности тренировок. И хотя МПК не увеличивается, у испытуемых продолжает улучшаться мышечная деятельность, требующая проявления выносливости. У них может повыситься способность работать при более высоком проценте МПК. Например, большинство бегунов могут пробежать дистанцию 42 км (26,2 мили) со средней скоростью, предполагающей использование 75 — 80 % их МПК [I].
Например, у бывшего рекордсмена мира в марафоне Альберто Салазара МПК составляло 70 мл-кг^-мин"'. Это ниже предполагаемого МПК на основании его результата 2 ч 8 мин. Однако он мог бежать дистанцию при 86 % МПК. Этот процентный показатель был намного выше, чем у других спортсменов. Именно этим можно частично объяснить его высокие спортивные результаты.
Главными показателями успешного выступления в видах спорта, требующих проявления выносливости, являются МПК и процент от МПК, который спортсмен способен поддерживать длительное время. Последнее, видимо, связано с порогом лактата, поскольку он, очевидно, является главным фактором, определяющим скорость, которую спортсмен способен выдерживать во время длительного соревнования. Таким образом, способность выполнять работу при более высоком проценте от МПК, скорее всего отражает более высокий уровень лактатного порога.
Так как индивидуальные потребности в энергии зависят от размеров тела, МПК, как правило, выражают относительно массы тела в миллилитрах кислорода, потребляемого на килограмм массы тела в минуту (мл-кг^-мин"'). Это позволяет более точно сравнивать индивидуумов разного размера, выступающих в таких видах, как бег. В то же время мышечная деятельность пловцов и велосипедистов более тесно связана с МПК, который выражают в литрах в минуту (л-мин~').
У физически активных учащихся колледжей в возрасте 18—22 лет средние показатели МПК составляют 38 — 42 мл-кг^-мин"' (девушки) и 44 — 50 мл-кг^-мин"' (юноши). Начиная с возраста 25 — 30 лет, у физически неактивных людей показатели МПК снижаются на 1 % в год. Очевидно, это обусловлено сочетанием двух факторов — биологического старения и малоподвижного образа жизни. Отметим также, что у женщин показатели МПК, как правило, намного уступают показателям мужчин одного с ними возраста. Это различие обусловлено двумя факторами: разным составом тела (у женщин, как правило, меньше чистая масса тела) и разной концентрацией гемоглобина (ниже у женщин, вследствие этого пониженная способность транспорта кислорода). Однако не совсем ясно, насколько это половое различие в МПК обусловлено действительно физиологическими различиями и насколько — традиционным малоподвижным
Уровни аэробной производительности порядка 80— 84 мл- кг~'-мин~1 характерны для сильнейших бегунов на длинные дистанции и лыжников. Самое высокое МП К у мужчин было зарегистрировано у чемпиона Норвегии по лыжным гонкам — 94 мл-кг~'-мин~'. Среди женщин самое высокое МПК зарегистрировано у лыжницы из России — 74мл-кг~'-мин~'. В то же время у недостаточно подготовленных людей МПК может быть ниже — 20 мл-кг~'-мин~'
образом жизни, который ведут женщины после достижения половой зрелости. Более подробно эти вопросы рассматриваются в главе 19.
ЭКОНОМИЯ УСИЛИЯ
По мере повышения эффективности выполнения какого-нибудь упражнения потребность в энергии снижается, и у вас повышается резерв-ность. Это иллюстрирует рис. 5.15 на основании данных двух бегунов на длинные дистанции. При всех скоростях выше 200 м-мин"' (7,5 миль-ч"'), бегун А потреблял значительно меньше кислорода, чем бегун Б. У обоих спортсменов были почти одинаковые МПК (64 — 65 мл-кг^-мин"'), поэтому бегун А, расходовавший меньшее количество энергии, имел очевидное преимущество во время соревнований.
Оба бегуна неоднократно принимали участие в соревнованиях. Во время соревнований по марафону они бегали со скоростью, требовавшей использования 85 % МПК. В среднем бегун А имел преимущество в 13 мин благодаря более высокой эффективности. Поскольку МПК у обоих спортсменов было одинаковым, а потребность в энергии разная, преимущество, которое имел бегун А, во многом было обусловлено более высокой эф-
102
фективностью его бега. К сожалению, мы не знаем, чем объясняются эти различия.
Исследования, проводившиеся на спринтерах, бегунах на средние и марафонские дистанции, показали, что наиболее экономичны и продуктивны марафонцы. Они, как правило, расходуют на 5 — 10 % меньше энергии, чем спринтеры и бегуны на средние дистанции. Правда, экономию усилий изучали только при относительно невысоких скоростях (10— 19 км-ч"', или 6—12 миль-ч~1). Вполне естественно предположить, что бегуны на длинные дистанции менее эффективны на коротких дистанциях по сравнению с теми, кто на них специализируется.
Различия в технике бега и специфичности тренировок в беге на короткие и длинные дистанции могут обусловливать указанные различия в экономичности. Как показывает анализ техники бега бегунов на средние, короткие и марафонские дистанции, у спринтеров и бегунов на средние дистанции значительно большее вертикальное движение при скорости бега 11—19 км-ч~1 (7 — 12 миль-ч"'), чем у марафонцев. Однако такие скорости значительно уступают соревновательной скорости бега на средние дистанции и поэтому, очевидно, неточно отражают эффективность бега спортсменов на более коротких дистанциях (1500 м и меньше).
Эффективность движения может оказывать еще большее влияние в других дисциплинах. Например, часть энергии, расходуемой во время плавания, используется для того, чтобы удержать тело на поверхности воды и произвести достаточное усилие для преодоления сопротивления движению воды. И хотя количество энергии, необходимой для плавания, зависит от размеров тела и "плавучести", главный показатель экономии плавания — эффективность приложения силы.
На рис. 5.16 показаны потребности в кислороде у тренированных пловцов обоего пола, а так-
Успех в видах спорта, требующих проявления выносливости, во многом зависит от
| Т |
• высокого МПК;
• высокого уровня лактатного порога или начала аккумуляции лактата в крови;
• большей экономии усилий или низкого показателя V'о, при выполнении работы одинаковой интенсивности;
• высокого процента медленносокращаю-щихся волокон
же у группы хорошо подготовленных троеборцев. Показатели потребления кислорода наносили на график при различных скоростях плавания. Хотя троеборцы ежедневно проводили тренировки по плаванию, ни один из них никогда не выступал в соревнованиях по плаванию. Любопытно, что хотя многие из троеборцев имели существенно более высокие показатели аэробной работоспособности, чем пловцы, лишь немногие из них могли демонстрировать такие же результаты, как наиболее слабый из пловцов. Некоторые пловчихи с МПК 2,1 — 2,3 л-мин"' проплывали дистанцию 400 м с таким же результатом, как и троеборцы с МПК выше 5,0 л-мин"'. Для пловцов была характерна более высокая эффективность.
Результаты во многих видах спорта могут в большей степени зависеть от техники спортсмена, чем от способности производить энергию. Поэтому важно не только направлять усилия на повышение выносливости и увеличение силы, но и на повышение технического мастерства спортсменов. Однако в видах спорта, требующих проявления выносливости, успех обусловлен, по крайней мере, тремя факторами:
• высоким показателем МПК;
• высоким порогом лактата или началом аккумуляции лактата в крови;
• большей экономией усилий или низким показателем V'о, при выполнении работы одинаковой интенсивности;
• высоким процентом медленносокращающих-ся мышечных волокон.
ЭНЕРГЕТИЧЕСКИЕ ЗАТРАТЫ ПРИ РАЗЛИЧНЫХ ВИДАХ ДЕЯТЕЛЬНОСТИ
Количество энергии, затрачиваемой при различных видах деятельности, зависит от интенсивности и вида физической нагрузки. Расход энергии при большинстве видов деятельности определяют по величине потребляемого кислорода, измеряя средний показатель потребления кислорода за единицу времени. На основании полученного показателя исчисляют количество килокалорий энергии, расходуемой ежеминутно (ккал-мин"').
Эти показатели обычно не учитывают анаэробные аспекты мышечной деятельности и из-
103
быточное потребление кислорода после выполнения работы. Это немаловажно, поскольку расход энергии при выполнении данной работы может составлять 300 ккал плюс дополнительные 100 ккал во время восстановления. Таким образом, общий расход энергии составит не 300, а 400 ккал.
Организму среднего человека необходимо 0,20 — 0,35 л кислорода в минуту для удовлетворения потребностей в энергии в состоянии покоя. Это составляет 1,0 — 1,8 ккал-мин"', или 60 — 108 ккал-ч^', или 1 44 —2 592 ккал-день"'. Очевидно, что любой вид деятельности увеличивает этот показатель. Диапазон суточного расхода энергии очень широк. Он зависит от многих факторов, в том числе от
• уровня двигательной активности;
• возраста;
• пола;
• размеров тела;
• массы тела;
• состава тела.
Также отличаются энергозатраты при занятии различными видами спорта. Некоторые виды спорта — стрельба из лука или игра в кегли — не намного превышают расход энергии по сравнению с показателем в состоянии покоя. Другим видам, таким, как спринтерские соревнования, требуется очень много энергии. Помимо интенсивности физической нагрузки, следует учитывать также ее продолжительность. Например, при беге со скоростью 25 км-ч"' (15,5 миль-ч~1) расходуется около 29 ккал-мин"', однако такую скорость можно сохранять лишь короткое время. В то же время при беге трусцой со скоростью 11 км-ч"' (7 миль-ч"') расходуется только 14,5 ккал-мин"', т.е. вдвое мень
ше. Однако скорость бега трусцой можно поддерживать значительно дольше, что приведет к большему общему расходу энергии.
В табл. 5.5 приведены средние показатели расхода энергии мужчин и женщин при различных видах мышечной деятельности. Большинство видов физической нагрузки, указанных в таблице, включают перемещение массы тела, поэтому приведенные значения могут значительно колебаться в зависимости от индивидуальных различий, а также уровня технического мастерства (эффективности движения).
В ОБЗОРЕ...
1. Скорость основного обмена представляет собой минимальное количество энергии, необходимой для осуществления основных клеточных функций. Ее измеряют с соблюдением определенных условий теста. Величина основного обмена тесно связана с чистой массой тела и площадью поверхности тела, хотя на нее могут влиять и другие факторы.
2. Величина основного обмена колеблется от 1 200 до 2 400 ккал-день"'. При выполнении повседневной мышечной деятельности расход энергии повышается до 1 800 — 3 000 ккал-день"'.
3. Метаболизм усиливается с увеличением интенсивности физической нагрузки, однако потребление кислорода ограничено. Его пиком является мпк.
4. Улучшение физической подготовленности часто означает более длительное выполнение работы при более высоком проценте МПК.
| Таблица 5.5 Расход энергии при различных видах физической деятельности |
| Вид деятельности | Мужчины, ккал-мин"' | Женщины, ккал'мин~' | Относительно массы тела, ккал-кг^-мин"1 |
| Баскетбол | 8.6 | 6,8 | 0,123 |
| Велосипедный спорт, миль-ч~' | |||
| 7,0 | 5,0 | 3,9 | 0,071 |
| 10,0 | 7,5 | 5,9 | 0,107 |
| Гандбол | 11,0 | 8,6 | 0,157 |
| Бег, миль-ч"' | |||
| 7,5 | 14,0 | 11,0 | 0,200 |
| 10,0 | 18,2 | 14,3 | 0,260 |
| Пребывание в положении сидя | 1,7 | 1,3 | 0,024 |
| Сон | 1,2 | 0,9 | 0,017 |
| Пребывание в положении стоя | 1,8 | 1,4 | 0,026 |
| Плавание (кроль, 3,0 мили-ч"') | 20,0 | 15,7 | 0,285 |
| Теннис | 7,1 | 5,5 | 0,101 |
| Ходьба, 3,5 мили-ч"' | 5,0 | 3,9 | 0,071 |
| Тяжелая атлетика | 8,2 | 6,4 | 0,117 |
| Борьба | 13,1 | 10,3 | 0,187 |
| Примечание. Показатели приведены для мужчин с массой тела 154 фунта (70 кг) и женщин с массой
| |||
| тела 121 фунт (55 кг). Они изменяются в зависимости от индивидуальных различий.
| |||
104
5. Улучшение работоспособности достигается также за счет повышения экономии усилий.
Истощение запасов гликогена
Поддержание уровней АТФ обеспечивается также аэробным и анаэробным расщеплением мышечного гликогена. В видах спорта, в которых интенсивные мышечные усилия длятся всего несколько секунд, мышечный гликоген является основным источником энергии для синтеза АТФ. К сожалению, его запасы ограничены и быстро истощаются.
Как и в случае с КФ, скорость истощения запасов мышечного гликогена зависит от интенсивности физической нагрузки. Увеличение интенсивности приводит к снижению уровня мышечного гликогена. Во время бега на спринтерские дистанции, например, мышечный гликоген расходуется в 35 — 40 раз быстрее, чем при ходьбе. Наличие мышечного гликогена может быть ограничивающим фактором даже при средних усилиях. Для удовлетворения высоких потребностей мышцы в энергии во время физической нагрузки необходимо постоянное поступление гликогена.
Как видно из рис. 5.17, мышечный гликоген быстрее используется в первые минуты физической деятельности. Несмотря на то, что испытуемый бежал в одном темпе, интенсивность исполь-
105
зования гликогена в икроножной мышце была максимальной в первые 90 мин. Испытуемый сообщал об испытываемом напряжении (как трудно ему давалось усилие) в различные моменты теста. Он испытывал среднюю степень стресса в самом начале, когда запасы гликогена были еще достаточно большими, несмотря на высокую интенсивность его использования. Он не испытывал значи
тельного утомления до тех пор, пока запасы мышечного гликогена почти не истощились. Таким образом, возникновение ощущения утомления при продолжительной физической нагрузке совпадает со снижением уровня мышечного гликогена. Марафонцы называют внезапное наступление утомления на 29— 35-м километре дистанции (18— 22-я миля) "столкновением со стенкой". Частично возникновение чувства утомления обусловлено истощением запасов мышечного гликогена.
Истощение запасов гликогена в различных мышечных волокнах. Существуют определенные паттерны рекруитирования вовлечения мышечных волокон и истощения в них запасов гликогена. Запасы гликогена могут истощиться в наиболее часто используемых волокнах. Это ведет к сокращению числа волокон, способных производить мышечное усилие, необходимое для выполнения физического упражнения.
На рис. 5.18 представлена микроструктура мышечных волокон, взятых из мышцы бегуна до и после забега на 30 км. На рис. 5.18,а мы видим дифференциацию МС- и БС-волокон. Одно из БС-волокон обведено кругом. На рис. 5.18,^продемонстрирован второй образец той же мышцы, окрашенной для выявления гликогена. Чем краснее (темнее) окраска, тем больше в ней гликогена. До забега все волокна содержали достаточное количество гликогена и имели красную окраску. В обведенном круге БС-волокне на рис. 5.18,6 (после забега) все еще достаточно гликогена. В то же время в находящихся рядом с ним МС-волок-нах запасы гликогена почти полностью истощены. Это свидетельствует о том, что МС-волокна более интенсивно используются во время мышечной деятельности, требующей проявления выносливости, во время которой производится среднее усилие, например, бег на 30 км.
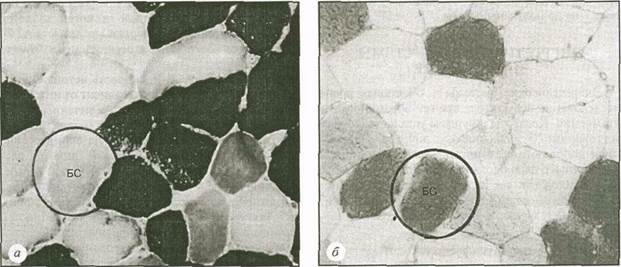
Рис. 5.18. Гистохимическое окрашивание на выявление мышечного гликогена до (а) и после (б)
забега на 30 км. Пояснения в тексте
106
Паттерн истощения запасов гликогена в МС-и БС-волокнах зависит от интенсивности физической нагрузки. Вспомним, что МС-волокна первыми вовлекаются при невысокой интенсивности физической нагрузки. По мере увеличения интенсивности начинают включаться и БСд-во-локна. Когда интенсивность возрастает до максимальной, наступает очередь БСц-волокон. По-видимому, такой же паттерн характерен и для процесса истощения запасов гликогена.
На рис. 5.19 показано количество гликогена, который использовали МС-, а также БС^- и БСд-волокна латеральной широкой мышцы бедра при езде на велосипеде с различным процентом МПК испытуемого. При относительно невысокой интенсивности физической нагрузки (40 — 60 % МПК) наиболее активными были МС-волокна. При более высокой интенсивности (75 — 90 % МПК) чаще использовались БС-волокна, которые быстрее истощали свои запасы гликогена по сравнению с МС-волокнами.
Это, однако, не означает, что МС-волокна меньше используются во время максимальных сокращений, чем БС, просто здесь отражен тот факт, что БС-волокна в большей степени "полагаются" на гликоген. При интенсивных мышечных сокращениях в работе участвуют все типы волокон.
Когда в МС-волокнах истощаются запасы гликогена, БС-волокна оказываются либо неспособными производить достаточное усилие, либо не могут быть полностью рекруитированы, чтобы компенсировать недостаточное усилие. Поэтому выдвигалось предположение, что ощущения мышечного утомления или тяжести при продолжительной физической нагрузке могут отражать неспособность некоторых мышечных волокон реагировать на требования, предъявляемые физической нагрузкой.
Истощение запасов гликогена в различных мышечных группах. Помимо избирательного истоще-
ния запасов гликогена в МС- и БС-волокнах, физические нагрузки могут предъявлять чрезмерно высокие требования к отдельным мышечным группам. Рассмотрим исследование, в котором испытуемые бегали на тредбане, установленном в трех положениях — горизонтально, с наклоном вниз и с наклоном вверх, — в течение 2 ч при 70 % МПК. Как показывают результаты (рис. 5.20), независимо от того, бежит ли спортсмен по наклонной вниз, вверх или горизонтально, больше всего гликогена использует икроножная мышца. Это свидетельствует о том, что мышцы-разгибатели голеностопного сустава, подвергавшиеся наибольшему физическому стрессу во время бега на длинные дистанции, скорее всего истощают запасы гликогена в отличие от мышц бедер. Таким образом, участком возникновения утомления являются мышцы нижней части ног.
107
коза, которая поступает в мышцы с кровью. Запасы гликогена в печени расщепляются, что обеспечивает постоянное поступление глюкозы в кровь. В первые моменты выполнения упражнения для образования энергии необходимо относительно немного глюкозы крови, однако позднее вклад глюкозы в образование энергии значительно увеличивается. Чтобы обеспечить адекватное потребление глюкозы мышцами, по мере увеличения продолжительности физической нагрузки печени приходится расщеплять все больше и больше гликогена.
Запасы гликогена в печени ограничены, и она не может быстро расщеплять глюкозу из других субстанций. Следовательно, когда потребление мышцами глюкозы превышает образование ее печенью, уровни глюкозы крови понижаются. Не имея возможности получить достаточное количество глюкозы из крови, мышцы вынуждены более интенсивно использовать свои резервы гликогена, что ускоряет истощение его запасов в мышцах и приводит к раннему возникновению утомления.
108
мышц. Многие ученые считают, что низкий мышечный рН является главным фактором, лимитирующим мышечную деятельность, а также основной причиной возникновения утомления при кратковременных максимальных физических нагрузках.
Как видно из рис. 5.21, восстановление после изнурительной физической нагрузки спринтерского характера уровня мышечного рН происходит в течение 30 — 35 мин. Даже при восстановившемся уровне рН концентрации лактата в крови и мышцах могут оставаться повышенными. Однако спортсмен может продолжать выполнять упражнение с относительно высокой интенсивностью даже при показателе мышечного рН ниже 7,0 и уровне лактата крови выше 6—7 ммоль-л"', т.е. в 4 — 5 раз превышающем уровень в состоянии покоя.
В последнее время некоторые тренеры и спортивные физиологи пытаются на основании измерения показателей лактата крови определить интенсивность и объем тренировок, обеспечивающих оптимальные тренировочные стимулы. Такие измерения позволяют определить интенсивность тренировок, однако они вряд ли могут охарактеризовать анаэробные процессы или степень ацидоза мышц. Поскольку лактат и Н+ образуются в мышцах, диффундируя из клеток во внеклеточные жидкости организма, затем транспорти-руясь к другим участкам тела для последующего обмена. Следовательно, показатели лактата крови зависят от интенсивности его образования, диффузии и окисления. На эти процессы может влиять множество факторов, поэтому целесообразность использования показателей лактата для определения объема и интенсивности тренировочных занятий несколько спорна.
НЕРВНО-МЫШЕЧНОЕ УТОМЛЕНИЕ
До настоящего времени мы рассматривали в пределах мышц факторы, которые могут являться причиной возникновения утомления. Однако ряд данных указывает, что при определенных обстоятельствах утомление может возникать вследствие неспособности активировать мышечные волокна. Это — функция нервной системы. Как рассматривалось в главе 3, нервный импульс передается по концевой пластинке двигательного нерва к мышце, чтобы активировать мышечную мембрану и заставить саркоплазматический ре-тикулум выделять кальций. В свою очередь, кальций, соединяясь с тропонином, инициирует мышечное сокращение. Обсудим два возможных нервных механизма, которые могут нарушить этот процесс и способствовать возникновению утомления.
Нервная передача
Утомление может возникнуть на концевой пластинке двигательного нерва мышцы, предотвра
щая передачу нервного импульса к мембране мышечного волокна. В исследованиях, проводившихся в начале столетия, наблюдали подобную неспособность передачи нервного импульса в утомленной мышце. Она может быть связана с одним или несколькими следующими процессами:
• выделением или синтезом ацетилхолина — нейромедиатора, переключающего нервный импульс с двигательного нерва на мембрану мышцы, количество которого может быть снижено;
• холинэстераза — фермент, расщепляющий ацетилхолин после того, как тот переключил импульс, может стать гиперактивной, предотвращая образование достаточной концентрации ацетилхолина для инициации потенциала действия;
• активность хорлинэстеразы может понизиться (гипоактивность), вследствие чего произойдет чрезмерное накопление ацетилхолина, парализующее волокно;
• может повыситься порог возбуждения мембраны мышечного волокна;
• некоторые субстанции могут вступить в борьбу с ацетилхолином за рецепторы на мембране, не активируя ее;
• калий может покинуть внутриклеточное пространство сокращающейся мышцы, снижая наполовину потенциал мембраны.
Большинство из этих причин нервно-мышечной блокады связаны с нервно-мышечными расстройствами (например, миастенией), но они также могут обусловливать некоторые формы нервно-мышечного утомления. Существует также предположение, что утомление может возникать в результате задержки кальция в Т-трубочках, что приводит к уменьшению количества кальция, необходимого для осуществления мышечного сокращения. В действительности истощение запасов КФ и накопление лактата могут просто увеличить интенсивность аккумуляции кальция в Т-трубочках. Эти теории возникновения утомления пока остаются лишь предположениями.
Центральная нервная система
Центральная нервная система (ЦНС) также может быть местом возникновения утомления. Имеются данные как подтверждающие, так и отрицающие это. Первые исследования показали, что когда мышцы испытуемого оказываются на грани крайней усталости, словесная поддержка, окрик или прямая электростимуляция мышц могут увеличить силу мышечных сокращений. Эти данные свидетельствуют о том, что лимитирующими факторами мышечной деятельности при выполнении изнурительной физической нагрузки могут быть и психологические факторы. Механизмы, лежащие в основе утомления ЦНС, недостаточно изучены. Неясно также, ограничена ли
109
эта форма усталости только ЦНС или же связана
с периферической передачей нервных импульсов.
Вовлечение в работу мышц частично зависит от сознательного управления. Психологическая травма изнурительной физической нагрузки может сознательно или несознательно подавить предрасположенность спортсмена выносить боль. ЦНС может снизить интенсивность нагрузки до терпимого уровня, чтобы "защитить" спортсмена. Ученые единодушны в том, что испытываемый дискомфорт утомления предшествует возникновению физиологических ограничений в мышцах. Большинство спортсменов, если только для них не характерен высокий уровень мотивации, прекращают выполнение упражнения, прежде чем наступит физиологическое изнеможение их мышц. Чтобы достичь пика мышечной деятельности, спортсмены должны на тренировках отрабатывать выполнение упражнений в наиболее оптимальном темпе, а также тренировать толерантность к утомлению.
В ОБЗОРЕ...
1. Утомление может возникать вследствие истощения запасов КФ или гликогена. В любом случае нарушается образование АТФ.
2. Очень часто в возникновении утомления "обвиняют" молочную кислоту. Действительно, образование молочной кислотой Н+ ведет к возникновению утомления. Аккумуляция Н+ снижает мышечный рН, что нарушает клеточные процессы образования энергии и сокращения мышц.
3. Возникновение некоторых форм утомления может быть обусловлено нарушением передачи нервных импульсов, в основе которого могут лежать различные механизмы.
4. ЦНС также может вызывать возникновение утомления, которое, по-видимому, играет защитную роль. Испытываемое утомление, как правило, предшествует физиологическому утомлению; утомленные спортсмены могут продолжить выполнение ----...„, „„^^„„^,1„ кш1у1 продолжить выполнение
упражнения после психологической поддержки.
В предыдущих главах мы рассмотрели совместные действия мышечной и нервной систем обеспечивающих движение. В этой главе мы обсудили метаболические процессы. Рассмотрели как хранится энергия в виде АТФ, как ее образование и наличие влияют на выполнение мышечной деятельности. Мы узнали, что метаболические потребности значительно изменяются. В следующей главе мы изучим управление метаболическими процессами, рассмотрев функции эндокринной системы.
Контрольные вопросы
1. Какова роль КФ?
2. Опишите взаимосвязь мышечных АТФ и КФ при спринтерской деятельности.
3. Почему систему АТФ — КФ и гликолитическую систему образования энергии считают анаэробными?
4. Какую роль играет кислород в процессе аэробного метаболизма?
5. Расскажите о промежуточных продуктах образования энергии за счет АТФ, гликолиза и окисления.
6. Что такое дыхательный коэффициент? Объясните его применение для определения окисления углеводов и жиров.
7. Какова взаимосвязь между потреблением кислорода и образованием энергии?
8. Что такое порог лактата?
9. Как на основании измерений потребления кислорода можно оценить производительность (эффективность) при выполнении упражнения?
10. Почему спортсмены с более высокими МПК показывают лучшие результаты в видах спорта, требующих проявления выносливости, по сравнению с теми, у которых эти показатели ниже?
11. Почему потребление кислорода часто выражают в миллилитрах кислорода на килограмм массы тела в минуту (мл-кг^-мин"')?
12. Опишите возможные причины утомления во время мышечной деятельности продолжительностью 15 — 30 с и 2 — 4 ч.
СПИСОК ЛИТЕРАТУРЫ
1. Со51И1 ^.^. (1970). Ме1аЬоИс гезропкек с1ипп§ сИагапсе гипгппе. .1оита1 оГАррНеД РЬу5ю1о§у, 28, 251 — 255.
2. Со51П1 ^.^., Оап;е1& .Г., Еуап<> \У., Рт1( V/., КгаЬепЬиЫ О., §аШп В. (1976). §1<е1е1а1 ти5с1е епгутез апД ПЬег сотроя1;юп т та1е апй Гета1е 1гас1( а1Ые1е8. 1оита1 оГАррНей РЬу5;о1оеу, 40, 149— 154.
3. СокНИ 0.1,., Ртк\У..1., Р1упп М., Юпуап ]. (1987). Ми&с1е ПЬег сотроэШоп апс1 епгуте ас1т11ех т еН1е Гета1е сН81апсе гиппегз. 1п1ета(юпа11оита1 оГ8рог(5 МесНсте, 8, 103 - 106.
4. СояИИ П.Ь- Рох Е.1- аШ\ . Р.пегееНсх оГ тагаИюп
—^ « ъ/«/ I (^у.
4. СохШ! О.Ь., Рох Е.Ь. (1969). Епе^еИсв огтагаПюп гиппше. МесЬсте апб 5с;епсе т ЗроПк, 1(2), 81—86.
5. Оо11п;с1< Р.О., Агтаи-опв К., 5аиЬеП С., Р;еЫ К 5а1пп В. (1972). Епгуте ас(т(у апс1 ЛЬег сотроаШоп т ^е1е(а1 ти5с1е оГ итгатес! апс1 1гатес1 теп. 1оита1 оГ АррЬес! Рпу5ю1оеу, 33, 312— 319.
6. 1уу .1.1.., \У;шеге К.Т., Уап Напс1е1 Р.;., Е1еег О Н СохпИ О.Ь. (1980). Ми5с1е ге5р;га(01у сарас;(у апс1 ПЬег (уре ах (1егеггп1пап15 оГ те 1ас1а1е (ЬгехЬоМ. 1оита1 оГ АррИей РЬу5ю1оёу, 48, 523 — 527.
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
Аппйгопе К.В. (1979). ВюсЬегтвПу: Епегеу КЬегаНоп апс1 и<>е. 1п К.Н.51гаи55 (Ей.), Зрот тесИсте апй рЬу51о1о8у. РЫ1ас1е1рЬ;а, РА: У/.В.Заипйеге
Вегёки-от }. (1967). Ьоса! сЬапеез оГ АТР ап<1 рЬохрЬосгеаИпе ;п Ьитап ти5с1е Из&ие т соппесИоп
110
ЭНЕРГИЯ ДЛЯ КЛЕТОЧНОЙ ДЕЯТЕЛЬНОСТИ
Источником всех видов энергии является энергия Солнца. Химические реакции, происходящие в растениях (фотосинтез), превращают энергию Солнца в химическую энергию. Употребляя в пищу овощи и фрукты, а также мясо животных, которые питаются растениями, мы получаем энергию. Энергия содержится в пищевых продуктах в виде углеводов, жиров и белков. Эти основные компоненты пищевых продуктов расщепляются в клетках нашего организма, освобождая энергию.
Поскольку энергия постепенно превращается в тепло, ее количество, высвобождаемое в результате биологических реакций, определяется по количеству образованного тепла. В биологических
85
системах энергия измеряется в килокалориях (ккал). Согласно определению, 1 ккал равна количеству тепловой энергии, необходимой для повышения температуры 1 кг воды с 1 до 15 °С. Например, при сжигании спички выделяется примерно 0,5 ккал, тогда как при полном сгорании 1 г углеводов выделяется 4,0 ккал.
Некоторое количество свободной энергии в клетках используется для развития и "ремонта" нашего организма. Такие процессы, как мы уже знаем, направлены на развитие мышечной массы под влияние тренировочных нагрузок и восстановление мышц после физических нагрузок или травм. Энергия также необходима для активного транспорта множества веществ, таких, как глюкоза и Са24' через клеточные мембраны. Активный транспорт необходим для функционирования клеток и поддержания гомеостаза. Определенное количество энергии используют миофибриллы для обеспечения скольжения филаментов актина и миозина, в результате которого производятся мышечное сокращение и сила, что мы уже рассматривали в главе 2.
ИСТОЧНИКИ ЭНЕРГИИ
Продукты питания состоят, в основном, из углерода, водорода, кислорода, а при наличии белков — из азота. Молекулярные связи в пищевых продуктах относительно слабые, и в случае расщепления освобождается небольшое количество энергии. Следовательно, продукты питания не используются непосредственно для потребностей клетки. Энергия молекулярных связей продуктов питания химически освобождается в клетках организма и хранится в виде высокоэнергетического соединения — аденозинтрифосфата (АТФ).
'у Образование АТФ позволяет клеткам со-т хранять энергию в этом высокоэнергети
Ческом соединении
В покое энергия, необходимая нашему организму, обеспечивается за счет расщепления практически одинакового количества углеводов и жиров. Белки представляют собой "строительные блоки" и обычно обеспечивают функционирование клеток небольшим количеством энергии. При увеличении мышечного усилия в качестве источника энергии больше используются углеводы. При максимальной кратковременной нагрузке АТФ почти исключительно образуется за счет углеводов.
Углеводы
Зависимость мышц от углеводов во время физической нагрузки связана с их наличием, а также способностью мышечной системы их расщеплять.
Содержание углеводов в печени и скелетных мышцах ограничено; их достаточно для образования не более 2 000 ккал энергии. Это количество расходуется на то, чтобы пробежать 32 км (20 миль). Запасы жиров достаточны для образования более 70 000 ккал энергии
Углеводы в конечном итоге превращаются в глюкозу — моносахарид, который транспортируется кровью ко всем тканям организма. В состоянии покоя поглощенные углеводы попадают в мышцы и печень, а затем преобразуются в более сложную молекулу сахара — гликоген. Гликоген находится в цитоплазме до тех пор, пока клетки не используют его для образования АТФ. Гликоген, содержащийся в печени, может снова превращаться в глюкозу, она транспортируется кровью к активным тканям, где и происходит ее метаболизм (расщепление).
Содержание гликогена в печени и мышцах ограничено, и его запасы могут истощиться, если в рационе питания нет достаточного количества углеводов. Таким образом, восполнение запаса углеводов во многом зависит от пищевых источников крахмалов и Сахаров. Без достаточного потребления углеводов мышцы и печень лишаются своего основного источника энергии.
Жиры
Жиры и белки также используются в качестве источников энергии. В организме содержится значительно больше жиров, чем углеводов. Как видно из табл. 5.1, жировой резерв энергии в организме значительно превышает углеводный. Однако жиры менее доступны клеточному метаболизму, поскольку прежде всего должна быть расщеплена сложная форма — триглицерид — на основные компоненты: глицерин и свободные жирные кислоты. Только свободные жирные кислоты используются для образования АТФ.
Таблица 5.1. Запасы "топлива" и энергии в организме
| Источник энергии | г | ккал |
| Углеводы гликоген печени мышечный гликоген глюкоза в жидкостях организма Всего Жиры подкожный внутримышечный Всего
| 110 250 15 |
0,451 1,025 0,062
1,538 70,980 1,465
|
| 375 7,800 0,161 | ||
| 7,961 72,445
| ||
| Примечание. Оценка произведена на основании средней массы тела 65 кг с содержанием жира 12 %.
| ||
86
Как видно из рис. 5.1, из одного и того же количества жиров и углеводов образуется абсолютно разное количество энергии: соответственно 9 и 4 ккалт'. В любом случае интенсивность освобождения энергии из этих соединений слишком небольшая, чтобы удовлетворить потребности организма в энергии во время интенсивной мышечной деятельности.
Белки
Процесс превращения белков или жиров в глюкозу называется глюконеогенезом. В результате серии реакций белок может превратиться в жирные кислоты. Это — липогенез.
Белки обеспечивают 5 — 10 % энергии, необходимой для выполнения продолжительного физического упражнения. Для образования энергии используются лишь основные единицы белка-аминокислоты.
ИНТЕНСИВНОСТЬ ВЫСВОБОЖДЕНИЯ ЭНЕРГИИ
Чтобы быть полезной, энергия должна высвобождаться из химических соединений с контролируемой интенсивностью. Частично эта интенсивность определяется выбором источника энергии. Если используется большое количество энергии из одного источника, клетки рассчитывают главным образом именно на этот источник. Такое влияние наличия энергии называется эффектом массового воздействия.
Специальные ферменты обеспечивают более четкий контроль интенсивности высвобождения свободной энергии. Многие из них облегчают расщепление (катаболизм) химических соединений (рис. 5.2). Хотя названия ферментов довольно сложны, все они заканчиваются суффиксом -аза. Например, фермент, воздействующий на АТФ, называется аденозинтрифосфатаза (АТФаза).
Разобравшись с источниками энергии, рассмотрим, как сохраняется энергия. В следующем параграфе мы изучим, как образуется содержащее энергию соединение — АТФ.
Жиры
(свободные жирные кислоты)
| Углеводы |
1 г С6Н12О6 1 г С„НзА
4 ккал энергии 9 ккал энергии
В ОБЗОРЕ...
1. Около 60 — 70 % энергии в организме человека превращается в тепло. Остальное количество используется для выполнения механической работы и осуществления клеточной деятельности.
2. Мы получаем энергию из пищевых продуктов — углеводов, жиров и белков.
3. Получаемая из продуктов питания энергия содержится в высокоэнергетическом соединении — АТФ.
4. Углеводы обеспечивают около 4 ккал энергии на 1 г, тогда как жиры —около 9 ккалт"1, однако энергия, получаемая из углеводного источника, более доступна. Белки также обеспечивают организм энергией.
Молекула АБ
| Фермент |
Рис. 5.1. Образование энергии из 1 г углеводов и ] г жиров
Фермент
Рис. 5.2. Действие ферментов в катаболизме (расщеплении) соединений
87
| Рис. 5.3 Структурный состав молекулы АТФ с энергобогатыми фосфатными соединениями (а) и высвобождение энергии (б) |
а ^АТФ^ = | Аденозин |— Энергия —(Р.)— Энергия —(Р.)—— Энергия —(Р.)
. АТФаза +
б | Аденозин~|-{р)—(Р}—(Р) ^—-^| Аденозин | - (Р)—(Р) + (р) + Энергия
АДФ
АТФ
БИОЭНЕРГЕТИКА: ОБРАЗОВАНИЕ АТФ
Молекула АТФ (рис. 5.3,а) состоит из аденози-на (молекулы аденина, соединенной с молекулой рибозы), соединенного с тремя группами неорганического фосфата (Р ). При воздействии фермента АТФазы последняя фосфатная группа отщепляется от молекулы АТФ, быстро высвобождая большое количество энергии (7,6 ккал-моль'' АТФ). В результате АТФ расщепляется на АДФ (аденозин-дифосфат) и фосфор (рис. 5.3,6). Однако где же находилась эта энергия в самом начале?
Процесс накопления энергии в результате образования АТФ из других химических источников называется фосфорилированием. Вследствие различных химических реакций фосфатная группа присоединяется к относительно низкоэнергетическому соединению — аденозиндифосфату, преобразуя его в аденозинтрифосфат. Когда эти реакции осуществляются без наличия кислорода, процесс называется анаэробным метаболизмом. Если же в реакции участвует кислород, процесс называется аэробным метаболизмом, а аэробное превращение АДФ в АТФ —окислительным фосфорилированием.
Клетки образуют АТФ с помощью трех систем: системы АТФ — КФ, гликолитической и окислительной систем.
СИСТЕМА АТФ - КФ
Простейшей энергетической системой является система АТФ — КФ. Кроме АТФ, клетки содержат еще одну богатую энергией фосфатную молекулу —креатинфосфат (КФ). Энергия, высвобождающаяся при расщеплении КФ, в отличие
от энергии, высвобождающейся при расщеплении АТФ, не используется непосредственно для выполнения работы на клеточном уровне. Она используется для ресинтеза АТФ, чтобы обеспечить его относительно постоянное образование.
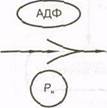
Высвобождению энергии при расщеплении КФ содействует фермент креатинкиназа, который действует на КФ с целью отделения Р^ от креатина. Освобожденная энергия может быть использована для присоединения Р^ к молекуле АДФ и образования АТФ (рис. 5.4). При использовании этой системы (энергия высвобождается из АТФ в результате отщепления фосфатной группы) клетки могут предотвратить истощение запасов АТФ, расщепляя КФ, и тем самым обеспечивая энергию для образования большего количества АТФ.
Это быстрый процесс, который может осуществляться без помощи каких-либо специальных структур клетки. Он может протекать и с участием кислорода, однако для его осуществления кислород не нужен, поэтому систему АТФ — КФ называют анаэробной.
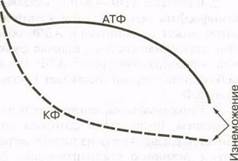
В первые секунды интенсивной мышечной деятельности количество АТФ поддерживается на относительно постоянном уровне, тогда как уровень КФ неуклонно снижается, поскольку он используется для пополнения запасов АТФ (рис. 5.5). В состоянии изнеможения уровни АТФ и КФ довольно низки и не могут обеспечить энергию для последующих сокращений и расслаблений мышц.
Таким образом, поддержание уровня АТФ за счет энергии, высвобождающейся при расщеплении КФ, ограничено. Запасы АТФ и КФ достаточны для удовлетворения энергетических потребностей мышц лишь в течение 3 — 15с спринтер-
|
|
| Энергия |
|
|
| ( аТФ) |
Рис. 5.4
Поддержание уровня АТФ за счет энергии,
содержащейся вКФ
88
| 100 |
| 80 |
| 60 |
| 40 |
| 20 |
2 4 6 8 10 12 14
Время,с
Рис. 5.5. Изменения в мышечных АТФ и КФ в первые секунды максимального мышечного усилия
ского бега. После этого мышцам приходится рассчитывать на другие процессы образования АТФ:
гликолитический и окислительный.
ГЛИКОЛИТИЧЕСКАЯ СИСТЕМА
Другой источник получения АТФ предусматривает высвобождение энергии в результате расщепления (лизиса) глюкозы. Это — гликолитичес-кая система, включающая процесс гликолиза, т.е. расщепление глюкозы с помощью специальных гликолитических ферментов (рис. 5.6).
Глюкоза составляет около 99 % всех Сахаров, циркулирующих в крови. Она поступает в кровь в результате усвоения углеводов и расщепления гликогена печени. Гликоген синтезируется из глюкозы вследствие процесса, называемого гликогенезом. Гликоген содержится в печени или мышцах до тех пор, пока не потребуется. Когда возникает потребность в гликогене, он расщепляется в результате процесса гликогенолиза на глюкозо-1 -фосфат.
Прежде чем глюкоза или гликоген могут быть использованы для образования энергии, они должны трансформироваться в соединение, которое называется глюкозо-6-фосфат. Для превращения молекулы глюкозы необходима одна молекула АТФ. При расщеплении гликогена глюкозо-6-фосфат образуется из глюкозо-1 -фосфата без затраты энергии. Гликолиз начинается, как только образуется глюкозо-6-фосфат.
Заканчивается гликолиз образованием пиро-виноградной кислоты. Для этого процесса не нужен кислород, однако использование кислорода определяет "судьбу" пировиноградной кислоты, образованной вследствие гликолиза. Когда мы говорим о гликолитической системе, мы имеем в виду, что процесс гликолиза протекает без участия кислорода. В этом случае пировиноградная кислота превращается в молочную кислоту.
Гликолиз, являясь более сложным процессом, чем система АТФ — КФ, обеспечивает расщеп
ление гликогена на молочную кислоту благодаря 12 ферментным реакциям. Все эти ферменты находятся в цитоплазме клеток. В результате гликолиза образуется 3 моля АТФ на каждый моль расщепленного гликогена. Если вместо гликогена используется глюкоза, образуется всего 2 моля АТФ, поскольку один моль расходуется на превращение глюкозы в глюкозо-6-фосфат.
Эта энергетическая система не обеспечивает образование большого количества АТФ. Несмотря на это, сочетанные действия гликолитической системы и системы АТФ — КФ обеспечивают производство силы мышцами даже при ограниченном поступлении кислорода. Эти две системы доминируют в первые минуты выполнения упражнений высокой интенсивности.
Другим значительным недостатком анаэробного гликолиза является то, что он вызывает накопление молочной кислоты в мышцах и жидкостях организма. В спринтерских дисциплинах продолжительностью 1 — 2 мин потребности гликолитической системы высоки, и уровни содержания молочной кислоты могут увеличиться с 1 (показатель в состоянии покоя) до более чем 25 ммоль-кг'. Такое подкисление мышечных волокон тормозит дальнейшее расщепление гликогена, поскольку нарушает функцию гликолитических ферментов. Кроме того, кислота снижает способность волокон связывать кальций и это может препятствовать сокращению мышц.
Интенсивность энергозатрат мышечного во-
Глюкоза Гликоген
Глюкозо-6-фосфат
Гликолитические ферменты
АТФ
Пировиноградная кислота
Молочная кислота
Рис. 5.6. Процесс гликолиза
99
Молочная кислота и лактат — не одно и то же соединение. Молочная кислота имеет формулу СдНдОд. Лактат представляет собой любую соль молочной кислоты. Когда молочная кислота теряет Н^ оставшееся соединение, соединяясь с Nа+ или К^ образует соль. В результате анаэробного гли-колиза образуется молочная кислота, которая очень быстро разлагается и образует соль — лактат. Из-за этого очень часто одно понятие используют вместо другого
локна во время нагрузки может быть в 200 раз выше, чем в состоянии покоя. Гликолитическая система и система АТФ — КФ не в состоянии обеспечить необходимое количество энергии. Без другой энергетической системы наша способность выполнять мышечную деятельность была бы ограничена всего несколькими минутами. Рассмотрим третью энергетическую систему.
ОКИСЛИТЕЛЬНАЯ СИСТЕМА
Последней системой образования энергии клеткой является окислительная система, наиболее сложная из трех энергетических систем. При ее рассмотрении мы опустим обременительные подробности. Процесс, в результате которого организм для производства энергии диссимилирует соединения, богатые энергией, с помощью кислорода называется клеточным дыханием. Это аэробный процесс, поскольку в нем участвует кислород. АТФ образуется в специальных клеточных органеллах — митохондриях. В мышцах они примыкают к миофибриллам, а также разбросаны по саркоплазме.
Мышцам необходимо постоянное обеспечение энергией для производства силы во время продолжительной мышечной деятельности. В отличие от анаэробного образования АТФ окислительная система производит значительное количество энергии, поэтому аэробный метаболизм — основной метод образования энергии во время мышечной деятельности, требующей проявления выносливости. Это предъявляет повышенные требования к системе транспорта кислорода к активным мышцам.
В ОБЗОРЕ...
1. Три энергетические системы обеспечивают образование АТФ:
• система АТФ — КФ;
• гликолитическая система;
• окислительная система.
2. В системе АТФ — КФ Р^ отделяется от кре-атинфосфата под действием креатинкиназы. Р^ затем может соединиться с АДФ, образуя АТФ. Это анаэробная система, главная функция которой — поддержание уровня АТФ. Величина высвобождения энергии составляет 1 моль АТФ на 1 моль КФ.
3. Гликолитическая система включает процесс гликолиза, посредством которого глюкоза или гликоген расщепляется на пировиноградную кислоту с помощью гликолитических ферментов. Если в процессе не участвует кислород, пирови-ноградная кислота превращается в молочную кислоту. Из 1 моля глюкозы образуется 2 моля АТФ, тогда как из 1 моля гликогена — 3 моля АТФ.
4. Гликолитическая система и система АТФ — КФ — основные источники энергии в первые минуты выполнения физического упражнения высокой интенсивности.
Окисление углеводов
Окислительное образование АТФ (рис. 5.7) включает три процесса:
1)гликолиз;
2) цикл Кребса;
3) цепочку переноса электронов.
Гликолиз при обмене углеводов играет важную роль как в анаэробном, так и аэробном образовании АТФ. Причем он протекает одинаково, независимо от того, участвует ли в процессе кислород. Участие кислорода определяет лишь "судьбу" конечного продукта — пировиноградной кислоты. Вспомним, что при анаэробном глико-лизе образуется молочная кислота и всего 3 моля АТФ на 1 моль гликогена. При участии кислорода пировиноградная кислота превращается в соединение, которое называется ацетил-кофермент А (ацетил-КоА).
Цикл Кребса. После образования ацетил-КоА попадает в цикл Кребса (цикл лимонной кислоты) — сложную последовательность химических реакций, которые позволяют завершить окисление ацетил-КоА. В конце цикла Кребса образуется 2 моля АТФ, а вещество (соединение, на которое воздействуют ферменты, в данном случае — первоначальный углевод) расщепляется и соединяясь с кислородом, образует диоксид углерода (СО-), который легко диффундирует из клеток, транспортируется кровью в легкие и выделяется во внешнюю среду (пост реакция декарбоксили-рования К - Н + СО,).
Цепочка переноса электронов. Во время гликолиза, когда глюкоза превращается в пировиноградную кислоту, выделяется водород. Значительно большее количество водорода выделяется во время цикла Кребса. Если он остается в системе,
90
| Рис. 5.7 Окисление углеводов |
| СНО (гликоген) |
| |
Глюкозо-6-фосфат
|
внутренняя часть клеток становится слишком кислой. Что происходит с этим водородом?
Таблица 5.2 Образование энергии вследствие окисления гликогена печени
| АТФ, образуемый из 1 моля
| ||
| гликогена печени
| ||
| Этап процесса | окислительным | |
| непосредственно | фосфорилированием* | |
| Гликолиз 3
| 6 | |
| (превращение
| ||
| глюкозы в пиро-
| ||
| виноградную
| ||
| кислоту)
| ||
| Превращение 0
| 6 | |
| пировиноградной
| ||
| кислоты
| ||
| в ацетил-КоА
| ||
| Цикл Кребса 2
| 22 | |
| 5
| 34 | |
| Итого 39
| ||
| * Относится к АТФ, образованному при переносе Н* И электронов в цепочку переноса электронов.
| ||
Цикл Кребса связан с серией реакций, которые называются цепочкой переноса электронов. Водород, выделяющийся во время гликолиза и в цикле Кребса, соединяется с двумя коферментами — ни-котинамид-аденин-динуклеотидом и флавин-аде-нин-динуклеотидом, которые переносят атомы во
дорода в цепочку переноса электронов, где они расщепляются на протоны и электроны. В конце цепочки Н"1" соединяется с кислородом, образуя воду и тем самым предотвращая подкисление.
Электроны, отделившиеся от водорода, принимают участие в серии реакций и в конечном итоге обеспечивают энергию для фосфорилиро-вания АДФ, а следовательно, образования АТФ. Поскольку этот процесс проходит с участием кислорода, он называется окислительным фосфорилированием.
Образование энергии за счет углеводов. Окислительная система образования энергии обеспечивает получение 39 молекул АТФ из одной молекулы глюкозы. Если процесс начинается с глюкозы, образуется 38 молекул АТФ (вспомним, что одна молекула АТФ используется до начала гликолиза для образования глюкозо-фосфата). В табл. 5.2 показано количество образующейся энергии.
Окисление жиров
Как уже отмечалось, жиры также вносят свой вклад в энергетические потребности мышц. Запасы гликогена в мышцах и печени могут обеспечить всего 1 200 — 2 000 ккал энергии, в то время как жиры, содержащиеся внутри мышечных волокон и в жировых клетках, — около 70 000 — 75 000 ккал.
Хотя жирами называются многие химические соединения, такие, как триглицериды, фосфоли-
91
|
|
пиды и холестерин, только триглицериды используются в качестве основного источника энергии. Триглицериды находятся в жировых клетках и волокнах скелетных мышц. Чтобы использовать триглицериды для образования энергии, необходимо расщепить их на основные составляющие:
одну молекулу глицерина и три молекулы свободных жирных кислот. Этот процесс называется липолизом и осуществляется ферментами — липазами. Поскольку свободные жирные кислоты — основной источник энергии, мы уделим им главное внимание.
Высвободившись из триглицерида (жира), свободные жирные кислоты могут попасть в кровь, которая транспортирует их по всему организму, и в результате диффузии проникнуть в мышечные волокна. Интенсивность поступления свободных жирных кислот в мышечные волокна зависит от градиента концентрации. Повышение концентрации свободных жирных кислот в крови выталкивает их в мышечные волокна.
B-Окисление. Несмотря на значительные структурные различия между разными свободными жирными кислотами, их метаболизм почти одинаков, как следует из рис. 5.8. До того, как они попадут в мышечные волокна, свободные жирные кислоты активируются энергией АТФ с помощью ферментов. Таким образом их подготавливают к катаболизму (расщеплению) в митохон-дриях. Этот ферментный катаболизм жиров митохондриями называется бета-окислением.
В этом процессе углеродная цепочка свободной жирной кислоты делится на двухуглеродные остатки уксусной кислоты. Например, если первоначально свободная жирная кислота имела 16-углеродную цепочку, то при (3-окислении образуется 8 молекул уксусной кислоты. Вся уксусная кислота затем превращается в ацетил-КоА.
Дата: 2018-12-28, просмотров: 394.