Важнейшими показателями динамики экосистем являются устойчивость и стабильность. Определение устойчивости экосистем уже давалось - это способность экосистемы возвращаться в исходное состояние после снятия внешнего воздействия, выведшего ее из равновесия. Под стабильностью экосистемы понимают ее способность сохранять свою структуру и функциональные свойства при воздействии на нее внешних факторов. Иногда понятия устойчивость и стабильность рассматриваются как синонимы, но тогда следует различать два вида устойчивости: резидентная устойчивость (стабильность) - способность оставаться в устойчивом (равновесном) состоянии под нагрузкой, и упругая устойчивость (собственно устойчивость) - способность быстро восстанавливаться при снятии нагрузки. В разных терминах имеются различные смысловые оттенки, которые нужно учитывать.
Системы с высокой резидентной устойчивостью способны воспринимать значительные воздействия, не изменяя существенно своей структуры, то есть практически не выходя за пределы равновесного состояния. Поэтому понятие упругой устойчивости для них не определено (если система не выходила за пределы равновесия, то как можно говорить о возвращении в равновесное состояние после снятия возмущения). Если внешнее воздействие превышает определенные критические значения, то такая система обычно разрушается. В технике подобное качество называется жесткостью. Предельные значения внешних воздействий, которые система способна выдержать без разрушения соответствуют запасу жесткости. Когда говорят о высокой резидентной устойчивости, то имеется в виду именно высокий запас жесткости данной системы. Это несколько отличается от понятия высокой стабильности, так как здесь в первую очередь внимание обращается на неизменность структуры. Тундра, например, обладает высокой стабильностью, но она очень ранима, у нее малый запас жесткости, то есть малая резидентная устойчивость. Экосистему тундры очень легко разрушить. Достаточно проехать вездеходу. Колеи, которые он оставляют за собой, сохраняются десятилетиями. Такие экосистемы по аналогии с техникой можно назвать хрупкими.
Системы с малой резидентной устойчивостью для нормального существования должны обладать высокой упругой устойчивостью. Они более чувствительны к внешним возмущениям, под действием которых они как бы “прогибаются”, частично деформируя свою структуру, но после снятия или ослабления внешних воздействий быстро возвращаются в исходное равновесное состояние. При превышении пороговых воздействий такая система теряет устойчивость, то есть все дальше удаляется от состояния равновесия. Диапазон воздействий, которые может выдержать система без разрушения, в технике соответствует запасу упругости. Таким образом, степень упругой устойчивости можно оценить как упругостью, определяющей степень сопротивления внешнему воздействию и скорость возврата в исходное состояние после снятия воздействия, так и запасом упругости. В отличие от упругих систем, пластичные системы после снятия внешнего воздействия не возвращаются в исходное состояние, а приходят к какому-то другому равновесному состоянию. Так согласно точке зрения оппонентов теории моноклимакса, для экосистем характерно не одно, а несколько состояний равновесия (климакса). Таким образом, для пластичных экосистем характерна малая упругая и малая резидентная устойчивость.
Похоже на то, что резидентная и упругая устойчивости взаимоисключают друг друга, точнее, экосистеме трудно развивать оба вида устойчивости. Например, одни леса состоят из деревьев с толстой корой, обладающих повышенной резидентной устойчивостью к пожарам. Но если такой лес все-таки сгорит, то его восстановление, как правило, крайне проблематично. Напротив, многие леса очень часто горят (низкая резидентная устойчивость), но быстро восстанавливаются (высокая упругая устойчивость). Ориентация экосистем на один из видов устойчивости определяется, как правило, изменчивостью среды: при стабильных условиях экосистемы склонны к более высокой резидентной устойчивости, при изменчивых условиях предпочтение отдается упругой устойчивости.
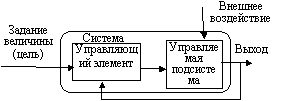
|
| Рис. 25 |
Природа обоих видов устойчивости состоит в наличии в экосистеме разного рода обратных связей. В основном это отрицательные обратные связи, которые направлены на стабилизацию параметров экосистемы, возвращая их значения к какой-то изначально заданной величине. Однако немаловажную роль играют и положительные обратные связи, усиливающие благоприятные для системы изменения, например, в плане роста и выживаемости организмов. Однако деятельность положительных обратных связей обязательно должна быть ограничена соответствующими отрицательными обратными связями, иначе ничем не контролируемая экспансия жизни может привести экосистему к гибели. В обоих случаях схема управления выглядит одинаково (рис.25).
Механизмы реализации обратных связей называются механизмами гомеостаза (от греческих слов homoios - подобный, одинаковый и ststis - состояние). Обычно они достаточно сложны, так как их компоненты связаны в единые информационные сети факторами среды, а также различными физическими и химическими агентами - “посредниками”, подобно тому, как нервная или гормональная системы связывают в одно целое части организма. Примером может служить сильная реакция организмов в экосистеме на слабые концентрации некоторых веществ. С энергетической точки зрения можно выделить одну закономерность, о которой уже говорилось ранее: обладатели высококачественной энергии, оставаясь в меньшинстве, управляют большими потоками энергии более низкого качества. Например, хищники управляют численностью травоядных животных. В то же время активность хищников и их численность управляется численностью их жертв по цепи обратной связи. По этой цепи на более высококачественный энергетический уровень (к хищникам) подается небольшая часть низкокачественной энергии - хищники уничтожают не всех жертв, а лишь небольшой их процент. Причем влияние этой части энергии на управление всей системой существенно усиливается деятельностью хищников.
Помимо отрицательной обратной связи устойчивость экосистемы может быть обеспечена избыточностью компонентов. Например, если в системе имеется несколько автотрофов, каждый из которых характеризуется своим температурным диапазоном функционирования, то скорость фотосинтеза сообщества в целом может оставаться неизменной, несмотря на колебания температуры. То же самое можно сказать и про гетеротрофов. Поэтому видовое разнообразие способствует повышению устойчивости экосистем (закон Эшби). При этом как в случае резидентной, так и в случае упругой устойчивости расширяется диапазон пороговых воздействий, которые способны выдержать экосистема без разрушения (запас жесткости и запас упругости).
Так, например, для тундры с ее бедным разнообразием характерны такие проявления низкой упругой устойчивости, как периодические резкие увеличения численности мелких грызунов - леммингов. О малой резидентной устойчивости тундры мы уже говорили. Экосистема тундры очень ранима и очень долго восстанавливается. В то же время тропические леса с их богатым разнообразием способны достаточно быстро залечивать свои раны. Легенды повествуют о целых городах, поглощенных джунглями, население которых не могло справиться с их натиском и вынуждено было уходить в другие районы.
Если разные стадии сукцессии экосистемы считать квазистатическими состояниями, то здесь можно отметить ряд закономерностей. На первых стадиях сукцессии экосистемы способны очень быстро возобновляться. Например, мы можем распахать по весне все поле , но уже к осени оно будет укрыто травами. Это говорит о достаточно высокой упругой устойчивости на начальных стадиях сукцессии, которая по мере движения к климаксу постоянно снижается. То есть на каждой следующей стадии для восстановления исходного состояния требуется гораздо больше времени. Климаксный лес характеризуется очень малой упругой устойчивостью.
Для резидентной устойчивости тенденция обратная. На первых стадиях сукцессии резидентная устойчивость остается достаточно невысокой - системы очень чувствительны к внешним воздействиям, откликаясь на них повышенным динамизмом. Например, заросли кустарника или молодой березняк может быть легко поврежден вездеходом (малая резидентная устойчивость), однако вторичная сукцессия достаточно быстро “затянет” повреждение (высокая упругая устойчивость). По мере роста видового разнообразия резидентная устойчивость непрерывно растет. При этом растет одновременно и жесткость системы (стабильность) и запас жесткости (взрослый лес вездеход так просто не одолеет). Однако по мере приближения к климаксу, как уже было показано, экосистема становится более чувствительна к действию некоторых катастрофических факторов среды (бури, пожары и т.п.). То есть на фоне продолжающегося роста стабильности (жесткости), запас жесткости экосистемы при движении ее к климаксу, по-видимому, начинает уменьшаться, что можно объяснить преобладанием в ней старых деревьев. Это говорит о том, что она становится все более хрупкой.
Как уже было сказано, для реальных лесов характерен, как правило, мозаичный климакс. По-видимому, именно такой лес имеет наилучший комплекс показателей устойчивости и стабильности. Периодические сильные воздействия на него в целом лес выдерживает достаточно жестко, жертвуя лишь наиболее старыми и хрупкими участками, которые, даже если они превращаются после пожара в абсолютно пустые пространства, в условиях непосредственной близости неповрежденного леса восстанавливаются сравнительно быстро.
Примером абсолютно нежизнеспособных экосистем являются агроценозы, создаваемы человеком из однолетних и двулетних культурных растений, то есть продукты сельскохозяйственной деятельности человека. Многие экологи даже не считают их экосистемами, хотя есть все основания считать их экосистемами с искусственно поддерживаемыми начальными стадиями сукцессии. Заброшенные поля сразу же втягиваются в естественный ход сукцессии. Что касается возделываемых полей, то они существуют только за счет человека. Свидетельством их нежизнеспособности является крайне малая устойчивость как упругая, так и резидентная, что является следствием очень скудного видового разнообразия как флоры (все сорняки подавляются гербицидами), так и фауны (“вредных” насекомых мы травим пестицидами). Малая упругая устойчивость проявляется, например, в резких вспышках численности конкретных видов вредителей. Подобные вспышки в естественной природе наблюдаются очень редко. Малая резидентная устойчивость проявляется, например, в повышенной чувствительности посевов к природным условиям, таким как град, засуха, повышенное количество осадков и т.д.
Осваивая все большие территории планеты, человек продолжает стратегию обеднения видового разнообразия. Этим он все более подрывает способность природы сопротивляться внешним воздействиям и возвращаться в исходное состояние. Как знать, может быть роковая черта, за которой начинаются необратимые изменения, уже пройдена. Это значит, что биосфера никогда больше не придет к исходному состоянию. Это значит, что мы вступили на путь глобальных перемен, которые постепенно изменят жизнь на Земле самым коренным образом. Будет ли в этом мире место человеку?
Глава 13.
Популяции в экосистеме
Цель: механизмы жизнедеятельности экосистем
Под популяцией можно понимать любую достаточно обособленную группу организмов одного вида, занимающую определенное пространство и функционирующую как часть экосистемы.
Различные виды организмов представлены в экосистеме именно популяциями. Поэтому популяцию можно рассматривать как систему, стоящую на иерархической лестнице природных систем между уровнем отдельных организмов и уровнем экосистем. Поэтому в организации популяций черты единого организма зачастую просматриваются более явно, чем в экосистемах. Правда, если экосистемы могут существовать относительно обособленно, как отдельные организмы, например экосистемы островов, то популяции в изолированном виде нигде не встречаются. Это своего рода функциональные органы экосистем, тесно взаимодействующие друг с другом, обеспечивая тем самым жизнедеятельность экосистемы. Хотя, по большому счету, и жизнь отдельных организмов лишена смысла вне тесной связи этих организмов друг с другом.
Популяция считается базовой единицей экологии. Если холистское направление в экологии на первое место ставит понятие экосистемы, рассматривая ее как единое целое, то редукционистское направление наибольшее внимание уделяет именно популяциям, считая что на этом уровне лежат все механизмы экосистемной динамики. Именно на уровне популяций происходят основные адаптации и эволюционные процессы, здесь рождается видовое многообразие природы, формируются межвидовые и внутривидовые взаимодействия и т.п.
Для человека знание законов популяционной динамики имеет одно из первостепенных значений, так как эти законы применимы и к динамике человеческих популяций. Особую важность эти знания приобретают в связи с демографической проблемой, являющейся одним из основных компонентов надвигающейся глобальной экологической катастрофы. Здесь же лежат ответы на вопросы о причинах нашей воинственности, стремления к безудержной экспансии планеты и даже космоса, наших конфликтов с природой. Распространяя законы популяционной динамики на человеческое общество, мы можем прогнозировать (имеется в виду именно научный прогноз, а не предсказание или гадание), что может ожидать нас в будущем. Правда, человек - это существо особое, в некоторой степени обособившееся от природы, обладающее мощными средствами воздействия на среду. Поэтому далеко идущие выводы могут быть абсолютно неправильными, как, например, неправильными оказались прогнозы многих поколений демографов прошлого, регулярно предсказывавших нам скорую стабилизацию численности населения планеты, чего до сих пор не произошло. Тем не менее основные тенденции мы представляем себе достаточно ясно. И выводы из анализа этих тенденций пока что говорят не в нашу пользу.
Динамика популяций
Прежде, чем говорить о динамике, рассмотрим ряд свойств, которыми характеризуется популяция. Эти свойства можно разделить на биологические и групповые. Биологические свойства присущи как популяции в целом, так и составляющим ее организмам. Они характеризуют жизненный цикл популяции: популяция, так же как и отдельный организм, растет, дифференцируется и поддерживает сама себя, имеет определенную организацию и структуру. Групповые свойства присущи только группе в целом. Как правило, это статистические параметры, такие как плотность, рождаемость, смертность, возрастная структура. В числе прочего, эти параметры хорошо характеризуют динамику популяции, то есть изменение во времени ее численности, структуры и т.п.
Плотность популяции выражается количеством особей или биомассой на единице пространства, например число деревьев на 1 га, или масса планктона в 1 м3. На разных стадиях жизненного цикла плотность может существенно колебаться. Это напрямую связано с двумя другими показателями популяции: рождаемостью и смертностью.
Рождаемость - это способность популяции к увеличению численности, независимо от того, происходит ли это путем откладывания яиц, или путем деления, или почкования, или прорастания из семени, или как-то иначе. Наиболее показательна удельная рождаемость, определяемая как количество особей, появившихся в единицу времени на одну особь в популяции (в демографии расчет ведется на одну женщину репродуктивного возраста). Реальная рождаемость в значительной степени зависит от факторов среды, поэтому она всегда меньше максимальной рождаемости, под которой понимают теоретический максимум рождаемости, определяемый только физиологией особей при оптимальных значениях всех факторов среды.
Смертность характеризует гибель особей в популяции и выражается числом особей, погибших в единицу времени. Удельная смертность характеризует уровень смертности для определенной группы особей, например, количество смертей за месяц на тысячу особей. Смертность также зависит от факторов среды и бывает обычно значительно выше минимальной смертности при идеальных условиях среды, которая определяется физиологией данного вида организмов - даже в идеальных условиях особи будут умирать от старости.
Выживаемость есть параметр, обратный смертности. Если смертность на N особей обозначить через М, то выживаемость определяется разностью N - M. Большой интерес представляют кривые выживания особей популяции в зависимости от их возраста.
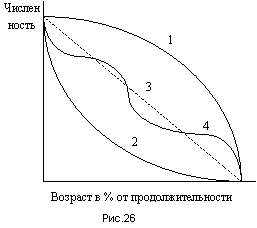
Различают три общих типа кривых выживания (рис.26). Выпуклая кривая 1 характеризует относительно низкую смертность на протяжении почти всего жизненного цикла популяции. К концу жизненного цикла смертность значительно возрастает. Такая кривая характерна для популяций, в которых основной акцент при размножении делается не на количество новорожденных особей, а на повышенную заботу о молодом поколении. Такого рода кривая характерна для многих высших животных, а также и для человека. Вогнутая кривая 2 характеризует повышенную смертность на ранних стадиях жизненного цикла, которая снижается, когда особи “входят в силу”, достигая определенного возраста. Такие кривые типичны для низших организмов и растений. Например, смертность среди молодых дубов и прорастающих желудей особенно велика, но когда дубы достигают определенных размеров, их смертность резко снижается. Аналогичные кривые характерны для рыб, где смертность среди мальков обычно гораздо выше, чем смертность среди взрослых особей. Повышенная смертность в раннем возрасте в таких случаях компенсируется повышенным количеством семян, икринок, яиц и т.п. Промежуточные типы кривой выживания колеблются обычно вокруг средней линии 3. Одним из примеров может служить ступенчатая кривая 4, характерная для видов, у которых смертность значительно меняется на разных стадиях жизненного цикла. Примером могут служить некоторые виды насекомых, жизненный цикл которых проходит последовательно через несколько стадий: яйцо, личинка, куколка, взрослое насекомое.
На основе кривых выживания можно уже сделать вывод о возрастной структуре популяции, которая определяет соотношение в ней разных возрастных групп. Знание возрастной структуры определяет возможность прогнозирования динамики численности популяции. Так, например, в популяциях, в которых ожидается быстрый рост численности, преобладают молодые особи. Такие популяции называют внедряющимися, или пионерными. Такое состояние популяции может быть вызвано освобождением экологических ниш (например, в связи с деятельностью человека), внедрением новых видов в экосистемы, где у них нет соответствующего хищника, причиной может быть также малая устойчивость экосистем и т.п. Если же в возрастной структуре достаточно велика доля старых особей, то в ближайшем будущем следует ожидать снижения численности популяции. Такие популяции называются регрессивными, или вымирающими. Наибольшей жизнеспособностью и стабильностью отличаются популяции, в которых возрастная структура более выровнена. Такие популяции называются нормальными.
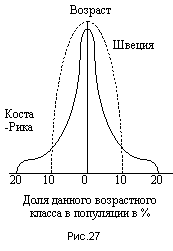
Особенно интересно приложение данной закономерности к человеческому обществу (рис.27). Сравнение возрастной структуры, например, в Швеции и Коста-Рике, красноречиво говорят об ожидаемой динамике численности в этих странах, что подтверждается фактическими данными: в Латинской Америке численность населения растет угрожающими темпами. Известно, что численность населения Латинской Америки по сравнению с 1900 годом возросла в 8,5 раз. В Европе ситуация более стабильная, однако наблюдается уменьшение численности коренного населения, некоторый прирост численности наблюдается лишь за счет эмигрантов.
Однако наибольшее значение для экологии имеют не сами показатели плотности или численности популяции, или ее возрастная структура, а скорости, с которыми меняются все эти показатели.
Если среда не лимитирует популяцию, то динамика численности популяции описывается дифференциальным уравнением: dN/dt = rN (скорость роста численности пропорциональна числу особей), которое имеет решение в виде: N = N0ert. Здесь N0 - численность в начальный момент времени, t - время, r - коэффициент роста популяции, определяемый как разность между удельной рождаемостью b (в расчете на одну особь) и удельной смертностью d в данный момент времени, то есть r = b - d. Максимально возможное значение коэффициента роста популяции rmax, достигаемое при наиболее благоприятных условиях среды, называется биотическим потенциалом популяции. Обычно биотический потенциал тем выше, чем ниже уровень организации организмов. Так дрожжевые клетки, размножаясь делением, при условии реализации биотического потенциала способны освоить весь земной шар за несколько часов. Крупным организмам с более низким потенциалом для этого потребовалось бы несколько десятилетий или даже столетий.
Следует отметить, что человек сейчас находится в условиях, когда его коэффициент роста близок к биотическому потенциалу. В 1968 году время удвоения численности людей составляло 35 лет, однако в некоторых странах этот период был еще меньше, и с течением времени он неуклонно уменьшается. То есть мы все ближе к некоторому минимальному значению периода удвоения численности, ниже которого мы не сможем переступить по чисто физиологическим причинам, это и будет, вероятно, соответствовать полной реализации нашего биотического потенциала. Понятно, что природа не выдержит такого натиска.
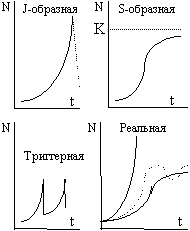
В соответствии с решением уравнения динамики численности популяции можно построить так называемую кривую роста популяции (рис.28). Из уравнения видно, что это экспонента, поэтому такую кривую называют экспоненциальной, или J-образной.
Особенность J-образной динамики в том, что рано или поздно относительно свободный рост численности популяции прекращается. Связано это, как правило, с исчерпанием какого-либо ресурса, на базе которого развивается данная популяция, например пространство или пища. Причиной также могут служить внезапные заморозки или какой-то другой фактор среды, прекращающий сезон относительного благополучия. После этого численность популяции может катастрофически понизиться (пунктирная линия на рисунке). Иногда вслед за подобным падением идет повторный рост численности, в результате формируется так называемая триггерная кривая роста, свойственная, например, однолетним растениям, некоторым насекомым, популяциям леммингов в тундре и т.п.
Подобные тенденции вызывают немалые опасения в плане судьбы человеческой цивилизации. Несмотря на все прогнозы демографов, динамика численности людей никак не желает стабилизироваться, оставаясь очень близкой к J-образной кривой. Мы уже близки к разрушению собственной среды обитания, а значит, вполне возможно, что в достаточно скором будущем (возможно, при жизни нынешних поколений) нас может ожидать катастрофическое снижение численности населения Земли.
Обычно лимитирующее давление среды играет в динамике популяции роль отрицательной обратной связи, поэтому в уравнении динамики популяции появляется дополнительный сомножитель или слагаемое: dN/dt = rN(1 - N/K) = rN - rN2/K, где К - верхняя асимптота, соответствующая верхнему пределу численности популяции после того, как в системе наступит равновесие (ее величина определяется средой, точнее, надсистемой). В данном случае скорость роста равна максимальной скорости роста (rN), определяемой биотическим потенциалом, умноженной на степень реализации максимальной скорости (1 - N/K), которая тем ниже, чем больше плотность, или численность N, популяции. Поэтому такую динамику называют зависимой от плотности (численности), в отличие от независимой от плотности J-образной динамики.
|
| Рис. 28 |
Решение этого дифференциального уравнения имеет вид: N = K/(1 + ea-rt), где a - постоянная интегрирования, определяющая положение кривой относительно начала координат. Кривая роста популяции, описываемая данным уравнением, называется логистической или S-образной кривой.
В принципе J-образную кривую можно рассматривать, как неполную реальную S-образную кривую, просто лимитирующие факторы среды в этом случае ограничивают рождаемость еще до того, как существенную роль в регулировке численности начнут играть внутренние факторы, обеспечивающие устойчивость системы. J-образная кривая роста характерна для некоторых популяций именно в системах с малой устойчивостью, то есть бедным видовым разнообразием, например, для леммингов в тундре, или для саранчи и других сельскохозяйственных вредителей в условиях агроценозов. Однако такая же независимая от плотности динамика характерна и для многих пионерных видов, заселяющих еще необжитые пространства или успешно вытесняющих более слабых конкурентов из их экологических ниш, как было, например, в случае с кроликами в Австралии. Нечто подобное, по-видимому, происходит и с человеческой цивилизацией.
Интересно, что оба основных типа роста численности популяции сходны с двумя типами метаболизма или роста для отдельных особей, что лишний раз подчеркивает глубокие аналогии между популяцией и живым организмом.
Почти все математические модели роста популяций сильно уязвимы. В наиболее частом случае динамика популяции не идет ни по одной из этих кривых, реальная кривая роста располагается где-то в промежутке между этими кривыми. По инерции рост численности еще некоторое время может продолжаться даже после выхода за пределы верхней асимптоты, но затем после нескольких колебаний численность популяции, как правило, либо стабилизируется, либо совершает незначительные колебания относительно значения N = K. Иногда эти колебания могут быть достаточно существенными, их называют популяционными волнами или волнами жизни. Причины популяционных волн могут быть как внешними (периодические изменения факторов среды), так и внутренними, связанными особенностями межвидовых и внутривидовых отношений.
Внутренние механизмы регулировки численности, работающие по принципу обратной связи, могут быть самыми различными. Все их можно объединить в понятие популяционного гомеостаза. Так наиболее действенным механизмом являются взаимоотношения между хищниками и жертвами, например рысь-заяц. Высокая численность жертв способствует росту хищников, которые сокращают поголовье жертв и сами попадают в неблагоприятные пищевые условия, в результате чего их численность начинает также уменьшаться, что способствует увеличению численности жертв, и т.д. Интересно, что статистическая модель системы хищник-жертва, реализованная на компьютере, дает колебания численности жертв и хищников, которые совершаются практически в противофазе, что вполне теоретически объяснимо, так как техническим аналогом данной биотической системы может служить обычный колебательный контур, где происходит периодическое взаимное превращение потенциальной и кинетической энергии. Однако для реальных кривых характерно лишь некоторое запаздывание по фазе колебаний численности хищников относительно колебаний численности жертв. Расхождение с моделью вызвано, по-видимому, наложением на систему хищник-жертва других механизмов гомеостаза, которые способствуют сближению фаз обоих колебаний, понижая тем самым инертность системы, а также тем фактом, что система эта не является изолированной, то есть заяц может стать жертвой совсем другого хищника, и рысь не обязательно питается только зайцами, имеются, вероятно, и другие причины. Система хищник-жертва может быть распространена на любой уровень трофической структуры экосистемы, например, заяц-трава и т.п.
К подобным межвидовым механизмам регулировки численности популяций можно отнести и межвидовую конкуренцию, которая ограничивает экологическую нишу данной популяции, приводя иногда даже к гибели части особей. При этом внутри популяции происходит достаточно жесткий отбор претендентов на продолжение рода. Конкретные механизмы этого отбора также могут быть различными. Это может быть по принципу “выживает сильнейший”, например, на стадии всходов молодых растений преимущество имеет тот, кто смог быстрее всех выйти в более высокий ярус, остальные погибают в затенении. Однако и они играют свою положительную роль в жизни популяции, отдавая, по-видимому, свою “жизненную силу” более перспективным претендентам через систему сросшихся друг с другом корней. У высших животных такая внутривидовая конкурентная борьба может проходить в более мягких формах, например, путем исключения ослабленных особей из процесса размножения без их непосредственной гибели.
Имеются и другие, менее понятные, механизмы. Например, зайцы-беляки в период пика численности могут часто погибать от “шоковой болезни”, связанной с увеличением надпочечников и другими признаками нарушения гормонального равновесия. Часто в условиях повышенной численности популяции у животных наблюдаются случаи каннибализма (поедания себе подобных), например окуни могут начать питаться собственными мальками. По мере возрастания биомассы деревьев более крупные и старые деревья могут становиться более восприимчивыми к насекомым-вредителям и часто гибнут, освобождая место более молодым. Иногда во время нашествий насекомых-вредителей такое массовое омоложение леса по масштабам может сравниваться даже с действием бурь или пожаров. Причем, в отличие от пожара, сукцессия начинается с более поздних стадий, а именно: со стадии молодых деревьев доминирующего вида. Интересным является механизм ингибирования (угнетения), проявляющийся в выделении в окружающую среду веществ, тормозящих динамику популяции. Этот механизм характерен как для растений и микроорганизмов, так и для животных. Например, подача в помещение, где нормально развиваются лабораторные животные (например, мыши), воздуха из густонаселенных помещений тормозит развитие животных в свободном помещении. Имеется и масса других примеров наличия внутренних механизмов регулировки численности. Практически все эти примеры можно свести к одному утверждению: в условиях повышенной плотности (численности) популяции возрастают стрессовые явления, которые действуют на данную популяцию угнетающе и являются своего рода внутрипопуляционной обратной связью.
В условиях человеческого общества, практически не имеющего врагов в природе, именно стрессовые механизмы начинают играть главную роль в ответ на взрывоподобный рост численности населения. Давно подмечено, что каждый человек обладает набором своего рода “интимных зон” разного радиуса, то есть определенных объемов пространства, где присутствие кого-либо вызывает неприятные ощущения. Так, например, в разговоре люди инстинктивно соблюдают определенную пространственную дистанцию между собой (порядка 1 м). Попытки сократить эту дистанцию вызывают у собеседника неосознанное желание чуть отодвинуться. Конечно, для разных людей эта зона различна. Имеются и другие зоны гораздо большего радиуса, но чем меньше радиус зоны, тем меньшее количество времени может выдерживать человек присутствие в этой зоне других людей без каких-либо стрессовых проявлений. Поэтому, например, люди, живущие в густонаселенных районах, в тесных комнатах многоквартирных домов, не имеющие возможности остаться наедине с собой в достаточно просторной зоне, оказываются гораздо более нервозными, чем, скажем, жители деревень, где такой проблемы не существует. Поэтому деревенские жители “задыхаются” в людных городах, поэтому горожан тянет “на природу”, где можно хотя бы на время расширить “зону интима”. Поэтому основное количество болезней (до 90 % и больше) сейчас либо напрямую вызвано нервными перегрузками, либо косвенно связано с нервным истощением организма. Так, может быть, механизм корректировки численности людей уже включился? Ведь не случайно, несмотря на успехи медицины, двадцатый век по праву можно назвать веком болезней.
Но все это лишь конкретные механизмы. Кто включает эти механизмы? Почему, например, после сильного выедания растительности леммингами в годы пика их численности происходит уменьшение доступности биогенных элементов, в результате чего ценность пищи леммингов снижается, в связи с чем ограничивается рост и выживаемость молодых особей? Затем биогенные элементы возвращаются в круговорот и растительный покров восстанавливается. Конечно, и здесь можно выявить определенные связи и адаптации. Однако ясно, что регулировка численности осуществляется не на популяционном, а на экосистемном уровне. То есть именно экосистема (надсистема по отношению к популяции) диктует популяции свои условия с помощью какой-то своей особой физиологии, что лишний раз доказывает ее аналогию с целостным организмом.
Вероятно, и человек в скором времени может испытать на себе прессинг надсистемы. Может быть, это уже происходит. Но слишком инертна надсистема, и слишком новым для нее является столкновение с человеком-разумным, вышедшим из-под власти биосферы. Когда она найдет действенное средство, ингибировать наш рост - лишь дело времени. Ни одна популяция, в том числе и человеческая не может жить вне экосистем, человек - часть организма планеты, и рано или поздно планета начнет “лечиться”.
Несколько слов нужно сказать и об одном из важнейших механизмов популяционного гомеостаза - миграции. Вообще переход особей из одной популяции в другую - дело обычное и полезное, так как способствует уменьшению вероятности близкородственного скрещивания. Правда, в периоды высокой плотности к иммигрантам относятся везде достаточно враждебно.
Однако наибольший интерес представляют массовые исходы особей из популяции при явной перенаселенности - нашествия. В популяциях, склонных к J-образной динамике (пионерные виды), нашествие является, по-видимому, одним из вполне обычных механизмов “деления” популяции, подкрепленных рядом инстинктов и адаптаций, в некотором роде подобных процессу деления клетки. Например, непосредственно перед миграцией у саранчи появляются более развитые крылья и более темная окраска, что способствует лучшему прогреванию тела солнечными лучами, а поэтому увеличивает подвижность особей. Сам процесс массовой миграции очень болезненный, так как гибнет огромное количество особей, у которых до некоторой степени притупляется инстинкт самосохранения. Все поведение насекомых направлено на выживание популяции, а не на личную безопасность.
У насекомых со сложной социальной организацией, типа пчел, муравьев и пр., подобные процессы настолько отлажены, что без особых преувеличений могут быть названы популяционной физиологией. Например, в пчелином улье в самом расцвете его сил, когда пчелиная семья наиболее сильна и запасы меда велики, рождается новая матка, а старая матка вместе с частью населения улья покидает его в поисках нового местожительства. Процесс этот происходит в несколько этапов, он настолько сложен и тонко “продуман”, что невольно напрашивается аналогия с процессом митоза (процесс деления эукариотических клеток). Я думаю, что процессы, подобные роению пчелиной семьи, являются более высокой эволюционной стадией того, что мы называем массовыми миграциями.
Не меньшее удивление вызывают периодические сезонные миграции птиц и рыб. Так, например, каждую осень мы наблюдаем, как многие виды птиц улетают на юг, а по весне возвращаются обратно. Механизм этих миграций, по-видимому, связан не с перенаселенностью, а с особенностями жизненного цикла, закрепленными в инстинктах. Однако некоторая общность все же имеется. Например, ближе к осени птицы становятся более беспокойными, усиливается стайный инстинкт, который дает эгрегору стаи (надсистеме) власть над каждой особью, заставляя ее, рискуя жизнью, пускаться в тяжелейшие испытания. Многие исследователи склоняются к мысли, что поведение птиц объяснимо только с холистских позиций. То есть главную направляющую роль в этих процессах играет биосфера в целом. Конечно речь не идет о каком-то разуме, командующем птичьими стаями. Однако в этих процессах можно наблюдать очень тесную аналогию с вегетативными процессами и системами организма (непроизвольные процессы, не подчиняющиеся непосредственно волевым усилиям, такие как сердечная деятельность, пищеварение, терморегуляция и т.п., в отличие, например, от двигательных систем).
Человек является, по-видимому, типичным пионерным видом с J-образной динамикой, для которого характерна тяга к миграциям в периоды перенаселенности. Для этого мы имеем определенный набор инстинктов. Так непосредственно перед миграцией у наших предков, по-видимому, начинали рождаться дети, для которых было характерно ускоренное созревание, более высокие физические данные и повышенная воинственность, потому что они должны были охранять мигрирующую популяцию, защищая ее ценой собственной жизни. По прибытии, они должны как можно скорее дать новое потомство, чтобы восполнить свои сильно поредевшие ряды. Со всеми этими атавизмами (признаки, свойственные далеким предкам) мы столкнулись, по-видимому, уже в наше время, что вызвало в свое время немалое удивление. Мы назвали это явление акселерацией (ускоренное развитие). Акселератам свойственны и быстрое половое созревание, и большая физическая сила, и повышанная агрессивность. В животном мире акселерация не редкость. Но возникает она как своего рода защитная реакция на грозящие данному виду потрясения. Возникает резонный вопрос: куда мы собираемся мигрировать? к каким потрясениям готовит нас эгрегор (надсистема) человеческого общества? А может он сам не понимает, что происходит и в беспорядке пробует все известные ему средства? Ведь мы живем не только во времена акселерации, но и во времена повышенного количества мутаций, вспомните, сколько детей с различного рода отклонениями рождается в наших роддомах. Что же нас ожидает?
Дата: 2018-11-18, просмотров: 1147.