Еще совсем недавно надежды человечества в плане решения продовольственной проблемы в связи с ростом народонаселения связывались с океаном, занимающим около 70 % поверхности Земли. Считалось, что море где-то раза в два плодороднее, чем суша. Там больше питательных веществ, растениям не нужны твердые ткани, чтобы тянуться к солнцу, и разветвленные корни, чтобы тянуться к влаге, более стабилен температурный режим и т.д. В океане очень велика скорость обновления живого вещества, так что растительность может давать до пятидесяти поколений в течение года. Поэтому, например, киты вырастают и становятся зрелыми не за 30-40 лет, как слоны, а всего за два года.
Казалось бы все логично. Однако более детальное исследование морских просторов, проводившееся 1964-1974 годах в рамках Международной биологической программы, показало, что море существенно уступает суше по средней продуктивности. Несмотря на свои масштабы, за год море дает всего 55 млрд тонн чистой первичной продукции, в то время как суша дает 115 млрд тонн в год.
Действительно некоторые зоны океана, особенно тропические прибрежные (шельфовые) зоны, особенно в местах впадения крупных рек (эстуарии), а также коралловые рифы и заросли водорослей отличаются наиболее высокой продуктивностью. Чистая первичная продуктивность эстуарий составляет 200-3500 в среднем 1500 г/см2 в год, продуктивность коралловых рифов и зарослей водорослей 500-4000 в среднем 2500 г/см2 в год. Такой же продуктивностью на суше обладают только влажные тропические и субтропические вечнозеленые леса (1000-3500 в среднем 2200 г/см2 в год). Это сравнимо с продуктивностью хорошо субсидируемых агроценозов, которая может достигать 3500 г/см2 в год (продуктивность несубсидируемых и малосубсидируемых агроценозров может опускаться до 100 г/см2 в год, в среднем эта величина составляет 650 г/см2 в год). Однако суммарная площадь эстуарий, рифов и зарослей водорослей очень мала и общая чистая продуктивность их составляет около 3,7 млрд т в год, в то время, как тропические и субтропические леса дают порядка 37,4 млрд т в год чистой продукции. Все остальные прибрежные воды характеризуются более низкими показателями первичной чистой продуктивности (200-600 в среднем 360 г/см2 в год). Однако в районах апвеллинга (плодородные районы океана с естественным притоком питательных веществ вследствие восходящего движения водных масс от дна к поверхности, вызванного, например, океаническими течениями) этот показатель возрастает (400-1000 в среднем 500 г/см2 в год).
Что же касается просторов открытого океана, то эти экосистемы характеризуются крайне низкой продуктивностью порядка 2-400 в среднем 125 г/см2 в год. Это почти в 2 раза ниже средней валовой первичной продуктивности по всей биосфере, которая составляет 333 г/см2 в год. Более низкий показатель 10-250 в среднем 90 г/см2 в год характерен только для пустынь и полупустынь. Даже тундре продуктивность несколько выше (10-400 в среднем 140 г/см2 в год). Для сравнения продуктивность лесов умеренной зоны составляет 600-2500 в среднем 1200 г/см2 в год, тайги - 400-2000 в среднем 800 г/см2 в год.
Столь низкая продуктивность океана объясняется тем, что основная масса воды океана очень бедна жизнью. Все продуценты обитают в верхнем слое воды. В прибрежных более плодородных зонах этот слой не превышает 30 м, в более бедных водах открытого океана зона продуцентов может простираться вниз на 100 м. Это связано с тем, что пик фотосинтеза приходится на самые верхние слои воды и его интенсивность быстро убывает с увеличением глубины. Основным продуцентом в море является фитопланктон, то есть мелкие водоросли, плавающие в толще воды, придающие ей зеленоватый цвет. Здесь же в богатых фитопланктоном поверхностных областях обитает и основная масса животных, для которых фитопланктон является основным поставщиком первичной продукции. Все эти организмы образуют так называемую поверхностную или планктонную пленку жизни.
Помимо поверхностной в океане принято выделять также донную, или бентосную пленку жизни, существующую в основном за счет оседания на дно отмершего вещества из поверхностной пленки. Ввиду невозможности фотосинтеза на больших глубинах эта пленка населена бентосом, то есть донными формами жизни, являющимися в основном гетеротрофами. Возможны также и другие типы глубоководных придонных экосистем, существующие за счет продукции хемосинтеза, связывающего энергию земных недр. Здесь помимо гетеротрофов имеются и автотрофы. Но эти экосистемы пока еще очень мало изучены.
Аналогичные пленки или сгущения жизни можно выделить и на суше. Это в первую очередь приземная пленка жизни, заключенная между поверхностью почвы и верхней границей растительного покрова. Это, собственно, та жизнь, с которой мы сталкиваемся ежедневно. Более скрыта от нас вторая, почвенная пленка жизни, населенная корнями растений, а также кротами, дождевыми червями, личинками насекомых и т.п., но основное население этой пленки составляют микроорганизмы. Несмотря на небольшую толщину, которая определяется в основном толщиной почвенного слоя, эта пленка характеризуется большим разнообразием жизни, чем приземная пленка. Достаточно сказать, что на 1м2 почвы можно насчитать миллионы насекомых, сотни дождевых червей, сотни миллионов микроорганизмов.
Впервые эти пленки, как наиболее важные очаги концентрации жизни, выделил В.И.Вернадский. Подобная пленочная организации жизни вовсе не случайна. Для биосферы вообще концентрация жизни подчиняется так называемому “краевому эффекту”: наибольшая концентрация жизни приурочена обычно к границам сред жизни. Это может быть поверхность океана или почвы, дно моря, а также и другие границы самого разного рода, например, опушка леса, пойма реки, берег моря и т.п. Общее название подобного рода границ - экотоны. Чем резче изменяется внешняя среда, тем более отчетливы границы сообществ по обе стороны экотона. Сообщества экотона содержат помимо организмов контактирующих сообществ еще и организмы, характерные только для экотона, которые за пределами экотона не встречаются. Дело в том, что многим видам организмов на разных стадиях жизненного цикла требуются сильно различающиеся условия, что наиболее удачно реализуется именно в экотонах. Например, многие личинки насекомых развиваются в воде, в то время как взрослые особи живут на суше; некоторым птицам нужны деревья для гнездовий и луга для питания и т.п. Поэтому зачастую число видов и плотность популяций в экотоне выше, чем в контактирующих сообществах. Особенно ярко краевой эффект проявляется на границах трех и более сред, например, дельта реки (река - море - суша - водух).
Человек всегда стремился селиться по берегам рек и морей. Если рядом был лес, то он стремился сохранить вокруг себя сообщество опушек, сокращая лес до небольших участков, перемежающихся с лугами и сельскохозяйственными угодьями. Это давало ему возможность пользоваться благами всех этих сообществ. Правда, всему есть предел. Например, чрезмерное увеличение периметра леса за счет увеличения количества опушек может привести к снижению продуктивности леса.
Среди наиболее типичных природных локальных сгущений жизни в океане, вызванных краевым эффектом, можно выделить следующие:
1. Прибрежные зоны океана.
Основная масса живого вещества в море сосредоточена около берега, где благоприятны условия питания. Именно в области континентального шельфа сосредоточено все крупное рыболовство мира. В прибрежной зоне можно выделить области литорали, то есть узкую зону прилива с периодически меняющейся средой. Особенно продуктивными областями прибрежной зоны являются эстуарии - системы, образуемые в местах впадения рек, протяженность которых тем значительней, чем больше вынос питательных веществ с суши.
2. Апвеллинговые зоны.
Процесс апвеллинга состоит в поднятии на поверхность холодной глубинной воды, богатой накопленными хорошо перемешанными питательными веществами. Обычно это происходит там, где ветры постоянно отгоняют поверхностную воду от крутого берегового склона. При этом в океане формируются особого рода циклонические течения, омывающие берега континентов. Эти зоны наиболее интенсивно используются для промысла рыбы. Интересно, что прибрежные районы суши из-за преобладания ветров, дующих в сторону моря и уносящих влагу с суши, представляют собой зачастую пустыни.
3. Коралловые рифы.
Система кораллового рифа формируется на основе некоторых водорослей и кишечнополостных животных, эволюционировавших совместно друг с другом. В результате возникает эффективный механизм круговорота элементов питания, позволяющий всей системе поддерживать очень высокую продуктивность в водах со сравнительно низким содержанием этих элементов. Обычно они формируются в районах с благоприятным температурным режимом. Они характеризуются огромным видовым разнообразием и богатством разного рода симбиозов, благодаря чему в них очень высок коэффициент эмерджентности, что и способствует одному из самых высоких показателей продуктивности.
4. Саргассовые сгущения.
Создаются большими массами плавающих водорослей, чаще всего саргассовых (в Саргассовом море) или филлофорных (в Черном море).
5. Рифтовые глубоководные сгущения.
Эти сгущения открыты только в 70-х годах. Они существуют в полной темноте в местах выхода горячей воды из разломов дна (рифтов), богатой минеральными солями и серой. Основным поставщиком первичной продукции для этих экосистем являются хемосинтезирующие серобактерии, высвобождающие энергию из соединений серы. Крупные многоклеточные организмы либо питаются этими бактериями, либо эти бактерии обитают у них в кишечнике, образуя уникальный симбиоз, характерный, по-видимому, для древнейшей жизни, развивавшейся за счет собственной энергии Земли. Об этой жизни мы знаем пока еще очень мало. Однако накопилось уже достаточно разного рода сенсационных сообщений, которые требуют тщательной проверки, о возможном наличии в этих экосистемах высоорганизованных форм жизни.
На суше также можно выделить ряд локальных сгущений жизни, связанных с краевым эффектом:
1. Экосистемы тропических и субтропических берегов морей.
Их высокая продуктивность связана с оптимальным достаточно стабильным температурным режимом и высокой влажностью. Особенно богаты эти экосистемы в случае, если от континентальной области их отделяют горы, не выпускающие тучи за пределы этих зон.
2. Экосистемы пойм рек, периодически заливаемые во время разливов.
Эти поймы хорошо удобряются илом, а вместе с ним органическими и биогенными веществами. Особенно продуктивны дельты рек, в которых из-за приносимого со всего водосборного бассейна вещества, в том числе и питательного, образуются целые острова.
3. Экосистемы небольших прудов и озер.
Особенно продуктивен пруд, расположенный, например, на краю лесной поляны. Наличие водоема способствует стабилизации баланса грунтовых вод в окрестности водоема, смягчает микроклимат, благотворно влияя на окружающую растительность. В то же время лес снабжает пруд питательными веществами (опавшие листья, сучья и т.п.). Поляна же обеспечивает наиболее благоприятные условия для существования животных, главным образом птиц и насекомых, личинки которых развиваются в пруду. Приходящие на водопой крупные животные удобряют поляну своими экскрементами.
Особо следует отметить экосистемы влажных тропических и субтропических лесов, расположенных в континентальной зоне. Хотя краевой эффект здесь проследить труднее, тем не менее они обладают продуктивностью, практически не уступающей продуктивности кораллового рифа. Секрет здесь тот же, что и в системах коралловых рифов, а именно: высокий коэффициент эмерджентности ввиду большого количества симбиотических связей. Другими словами, тропический лес представляет собой отлаженный организм, способный существовать даже в условиях бедности свежих поступлений питательных веществ, жизнь которого мало зависит от “превратностей” внешней среды. Как в любом живом организме, в нем организованы внутренние круговороты веществ, уникальные цепи преобразования и передачи энергии. Если однажды разрушить все эти взаимосвязи, то такая система не сможет заново воссоздать себя, даже если мы предоставим для этого все необходимые компоненты, так же как невозможно создать живую клетку собрав воедино все то, из чего она должна состоять, так же как невозможно осуществить опыт доктора Франкенштейнга (создать живого человека из фрагментов различных трупов).
Экологические пирамиды

|
| Рис. 21 |
Трофическую структуру экосистемы можно изобразить в виде разного рода экологических пирамид (рис.21).
Наиболее показательна пирамида энергии, которая наглядно иллюстрирует “правило десяти процентов”. Она строится из прямоугольников, поставленных друг на друга. Длина основания каждого прямоугольника в определенном масштабе соответствует величине потока энергии на разных уровнях трофической системы, точнее, той части энергетического потока, которая идет на формирование биомассы данного уровня, то есть на продукцию. Поэтому пирамида энергии, построенная в единицах измерения продуктивности, называется пирамидой продуктивности. Основанием пирамиды служит прямоугольник, соответствующий уровню продуцентов.
Важность пирамиды энергии определяется тем фактом, что число и масса организмов на данном трофическом уровне зависит не от количества биомассы (накопленной энергии) на предыдущем уровне, а от скорости продуцирования этой биомассы, являющейся пищей для данного уровня. Пирамида энергии (продуктивности) как раз и отражает картину скоростей прохождения массы пищи через пищевую цепь.
Как уже говорилось, из всей энергии, потребляемой организмами данного трофического уровня, в среднем только 10 % накапливается в биомассе, то есть идет на создание продукции данного уровня и становится доступной для консументов следующего трофического уровня. Из этой энергии опять же только около 10 % расходуется на вторичную продукцию. Поэтому продуктивность каждого следующего уровня где-то в 10 раз меньше продуктивности предыдущего уровня. Это напрямую вытекает из второго закона термодинамики и принципа Онзагера: повысить качество определенной порции энергии можно только путем соответствующего преобразования гораздо большего количества менее качественной энергии.
Поэтому пирамида энергии (продуктивности) любой экосистемы всегда имеет правильную форму, то есть подчиняется правилу: на каждом последующем уровне пищевой цепи продуктивность меньше, чем на предыдущем. Более того, суммарное количество вторичной продукции, образующейся на разных трофических уровнях, меньше первичной продукции данной экосистемы.
Пирамида биомассы строится аналогичным образом. Она характеризует общую массу живого вещества в сухом весе на каждом трофическом уровне экосистемы. Такая пирамида хорошо характеризует трофическую структуру экосистемы для оценок урожаев на корню. Для разных экосистем эта пирамида может выглядеть по-разному. Пирамиды биомассы экосистем суши обычно характеризуются такой же формой, как и пирамида энергии. То есть количество биомассы на каждом последующем уровне такой экосистемы как правило меньше, чем на предыдущем уровне. Но это правило уже не является абсолютным. Так для экосистем моря как правило характерна перевернутая пирамида биомассы, основание которой меньше, чем последующие ступени. Так общая биомасса всех потребителей фитопланктона может быть существенно выше, чем масса самого фитопланктона, совокупная масса крупных рыб может оказаться меньше массы мелких рыб. Такая ситуация вообще характерна для экосистем с очень мелкими продуцентами и крупными консументами. Причина этого - резкие различия в продолжительности жизни и продуктивности организмов на разных уровнях. Например, время жизни фитопланктона оценивается несколькими днями или даже часами, в то время как крупные животные могут десятилетиями накапливать массу. В то же время фитопланктон имеет большую продуктивность, но вся продукция достаточно быстро выедается, так что урожай на корню фитопланктона в каждый момент времени оказывается сравнительно малым. При всем этом через трофический уровень продуцентов проходить гораздо больший поток энергии, чем через уровни консументов. То есть пирамида энергии имеет обычную форму.
Численность организмов на каждом трофическом уровне экосистемы также можно изобразить в виде пирамиды численностей. Однако такая пирамида не очень показательна. Ввиду существенного разброса численности организмов на разных уровнях ее трудно изобразить в одном масштабе. Форма ее может быть самая различная. Например, для морских экосистем основание пирамиды должно отражать огромное количество организмов фитопланктона. Луговые продуценты (травы) также достаточно многочисленны, по крайней мере в сравнении с количеством продуцентов леса, представленных главным образом деревьями и кустарниками. Основной недостаток пирамиды численностей в том, что на одном трофическом уровне могут находиться особи, существенно различающиеся по размерам. Однако в любом случае эта пирамида свидетельствует о том, что количество организмов уменьшается от основания к вершине. Правда, это правило не относится к детритным пищевым цепям, то есть к редуцентам, представленным главным образом микрооорганизмами.
Разложение живого вещества
Разложение есть результат биотических (связанных с деятельностью живых организмов), так и абиотических (непосредственно с жизнью несвязанных) процессов. Типичным абиотическим процессом разложения биоорганики является пожар. Пожар - это важный, а иногда и необходимый процесс в экосистемах, где организмы-деструкторы не успевают возвращать вещество в круговорот жизни. Например, сильные (верховые) лесные пожары оставляют после себя удобренную почву, на которой через сравнительно короткое время разовьется более молодой лес. К абиотическим деструкторам можно отнести также процессы замораживания и оттаивания, трение об воду и т.п.
Однако главную роль в процессах разложения играют живые существа. Все консументы, начиная с фитофагов и кончая сапротрофами, участвуют в процессе измельчения и переваривания пищи. То есть процесс разложения происходит в основном благодаря преобразованию энергии внутри организмов и передаче ее от одного организма к другому вдоль пищевой цепи. Этот процесс необходим для жизни, и в случае его прекращения вскоре все биогенные элементы оказались бы сосредоточенными в мертвых остатках, после чего жизнь прекратилась бы. В природе процесс разрушения примерно уравновешивает первичную продуктивность.
Основную работу в процессах распада биоорганики выполняют сапротрофы, в основном грибы и бактерии. Однако присутствие животных, особенно мелких (короеды, улитки, жуки-могильщики и т.п.) существенно ускоряет процесс разложения. Например животные-детритофаги не могут переварить целлюлозу, но они способны измельчить ее, увеличивая площадь, доступную для воздействия микроорганизмов. Кроме того, они выводят в среду белки, стимулирующие рост и активность микроорганизмов. Одновременно они выедают часть бактерий и грибов, то есть участвуют в регулировании всего детритного комплекса. Процесс разложения довольно сложен и детально отлажен (так и хочется сказать “продуман”). Ни один вид сапротрофов не может самостоятельно осуществить разложение отмерших остатков. Полное разрушение осуществляется целым комплексом разрушителей, которые последовательно сменяют друг друга, подготавливая среду друг для друга.
В процессе разложения можно выделить три стадии:
1) размельчение детрита, сопровождающееся высвобождением растворимого органического вещества;
2) сравнительно быстрое образование гумуса и высвобождение сапротрофами дополнительного количества растворимых органических веществ:
3) более медленная минерализация гумуса.
Гумус с химической точки зрения представляет собой комплекс продуктов конденсации ароматических соединений (фенолов) с продуктами распада белков и полисахаридов. Детрит (продукты распада отмершей органики), гумус и другой органический материал играют важную роль при образовании почв. Они придают почве структуру, благоприятную для роста растений, облегчают усвоение растениями питательных веществ.
Минерализация органических веществ, в результате чего растения снабжаются минеральным сырьем, является не единственной функцией сапротрофов. Как уже говорилось, они могут служить пищей для некоторых животных (например, грибы, собираемые нами в лесу). Кроме того, грибы и микробы, несмотря на то, что мы считаем их примитивными, быстро приспосабливаясь к меняющимся условиям, обеспечивают “тонкую настройку”, поддерживающую стабильность экосистемы. Они выделяют в окружающую среду некоторые вещества (энзимы), оказывающие управляющее действие на всю экосистему, стимулируя или подавляя (ингибируя) активность отдельных подсистем. Это в первую очередь “наружные гормоны” или “гормоны среды”, например пенициллин, выделяемый плесневым грибом, подавляющий деятельность бактерий, а также разного рода витамины, стимулирующие их рост. Таким образом, благодаря сапротрофам, почва, как и живая ткань, обладает богатым набором ферментов и проявляет каталитическую активность. В ней протекают сложнейшие процессы обмена веществ и энергии, идет непрерывное производство определенных органических веществ и переход сложных соединений в более простые, доступные растениям. В почве, как в любом живом организме, все сложнейшие процессы происходят с явными признаками упорядоченности. Все это позволяет рассматривать почву со всеми населяющими ее микро- и макроорганизмами, как единое биологическое тело, то есть живое существо.
Для биосферы важное значение имеет некоторое отставание разложения продукции автотрофного уровня от процессов ее создания, благодаря чему стабилизируется круговорот биогенных веществ и состав атмосферы (соотношение СО2 и О2), а также происходит накопление горючих ископаемых. Поэтому крайнюю озабоченность вызывает деятельность людей, которые ускоряют процессы разложения сжигая горючие ископаемые, разрушая структуру почвы, уничтожая леса. В результате количество углекислого газа в атмосфере неуклонно возрастает, что заставляет нас беспокоиться по поводу грозящего планете потепления. В отличие от колебаний средней температуры планеты в прошлые эпохи, это потепление развивается стремительными темпами. Мы уже подняли среднюю температуру планеты где-то на 2 градуса по сравнению с доиндустриальной эпохой. Еще на 2 градуса выше температура будет уже в 2050 году, если к тому времени цивилизация еще будет существовать в современном виде. Этот процесс рушит многие механизмы устойчивости экосистем. Невооруженным глазом видно, как природа уступает стремительному натиску человека. Сможем ли мы сами выжить в стерильном мире? Конечно же, нет. Жизнь процветает благодаря разнообразию и тесному переплетению различных форм.
Может быть, поэтому природа активизирует против нас мир сапротрофов, поражая нас все новыми болезнями. Ведь сапротрофы пожирают не только отмершие организмы, но и ослабленные. А ослабление наших организмов вызвано тем образом жизни, который дисгармонирует с природой. Микроорганизмы очень динамичны и чувствительны к малейшим изменениям среды, а может, и к изменениям биополя планеты, выполняя роль гормонов, управляющих всеми процессами в биосфере. В определенном смысле именно они являются “царями природы”, корректируя численности популяций макроорганизмов. Не это ли “голос планеты”, все более дающий о себе знать в ответ на наше давление на нее?
Глава 10.
Среда обитания
Цель: факторы среды и адаптация к ним организмов
Влияет ли планета на жизнь в биосфере посредством планетарного биополя или нет - вопрос спорный. Однако ни у кого не вызывает сомнения то, что сама среда, в которой существуют организмы, оказывает на них регулирующее действие. Природные условия в различных районах планеты могут существенно различаться. В то же время жизнь обладает удивительным свойством адаптироваться к изменениям окружающей среды. И эти адаптации даже в удаленных и не связанных друг с другом районах, но с аналогичными условиями среды оказываются удивительно похожими, в результате чего там развиваются похожие формы жизни. Тем самым планета, а также Солнце, Луна, другие планеты и вообще весь космос как бы направляют эволюционные процессы в биосфере. Сама структура планеты, ее ландшафты, ее место в солнечной системе и во Вселенной в целом в совокупности с пройденным историческим путем (памятью) несут в себе в “закодированном” виде информацию о возможных формах жизни в этих условиях. Именно этот код и пытается разгадать наука в поисках закономерностей глобального эволюционного процесса. Может быть, именно это является своего рода генетическая программой, определяющей эволюцию планеты в данный момент времени. Тем более, что в самих этих внешних для жизни факторах заложена определенная фрактальная повторяемость (как и в структуре ДНК), в основе которой лежат законы Вселенной.
Пытаясь понять непонятное, человек жаждет чуда. Для решения таинственных загадок он изобретает полумистические гипотезы, которые, кстати сказать, не обязательно являются заблуждениями, достаточно вспомнить парадоксы квантовой механики или теории относительности. Однако многие чудеса, действительно существуя в реальности, имеют на удивление простые механизмы. Например, вавилонское чудо, когда Бог смешал языки людей, после чего они перестали понимать друг друга, мы наблюдаем сейчас в науке, где огромное количество узких отраслей знания создали свою уникальные терминологии, облегчающие общение между специалистами в данной отрасли, но затрудняющие межотраслевое общение. Может быть, то же самое произошло и с древними архитекторами, землемерами, каменщиками, столярами и др., совместно участвующими в построении грандиозного сооружения, переставших понимать друг друга, а потому перессорившимися и разбежавшимися в разные стороны, дав начало отдельным народам. Ведь не случайно разные народы имеют свои характерные особенности: есть народы-торговцы, народы-строители, народы-землепашцы, народы-воины, народы-мыслители.
Возможно, теория биополя также найдет когда-нибудь простое и ясное толкование, после чего обнаружится простота действующих при этом механизмов. На мой взгляд, достаточно большую роль в этом толковании будет играть концепция среды обитания.
Понятие среды обитания
Под средой обитания понимают совокупность внешних природных условий и явлений, в которые погружены живые организмы, и с которыми эти организмы находятся в постоянном взаимодействии.
В биоэкологии речь обычно идет о природной среде, не измененной человеком. В прикладной (социальной) экологии говорят об окружающей среде, так или иначе опосредованной человеком.
Отдельные элементы среды обитания, на которые организмы реагируют приспособительными реакциями (адаптациями), называются экологическими факторами или факторами среды. Среди факторов среды выделяют обычно три группы факторов: абиотические, биотические и антропогенные.
1. Абиотическими факторами среды называются условия, напрямую не связанные с жизнедеятельностью организмов. К числу наиболее важных абиотических факторов можно отнести температуру, свет, воду, состав атмосферных газов, структуру почвы, состав биогенных элементов в ней, рельеф местности и т.п. Эти факторы могут воздействовать на организмы как непосредственно, например свет или тепло, так и косвенно, например рельеф местности, обусловливающий действие прямых факторов, света , ветра, влаги и пр. Возможно, среди абиотических факторов присутствуют и такие , о которых мы пока даже не догадываемся. Так, например, мы совсем недавно открыли влияние изменений солнечной активности на процессы в биосфере.
2. Биотическими факторами среды называется совокупность влияний одних организмов на другие. Живые существа могут служить источником пищи для других организмов, являться средой их обитания, способствовать их размножению и т.п. Действие биотических факторов может быть не только непосредственным, но и косвенным, выражаясь в корректировке абиотических факторов, например, изменение состава почвы, микроклимата под пологом леса и т.п.
3. Антропогенными факторами среды называется совокупность влияний человека на живые организмы. Это влияние также может быть прямым, например, когда человек вырубает лес или отстреливает животных, или косвенным, проявляющимся в воздействии человека на абиотические и биотические факторы среды, например, изменение состава атмосферы, почвы, гидросферы, или изменение структуры экосистем.
Существование любого организма зависит от целого комплекса факторов. При этом удается выделить ряд закономерностей, общих для самых различных частных случаев.
Так изучая действие факторов среды на рост растений, в 1840 г. Ю.Либих установил, что урожайность различных культур определяется не теми веществами, которые присутствуют в относительном изобилии в окружающей среде, например, углекислый газ, или вода, а элементами, жизненно важными, но присутствующими в среде в малых количествах, например, цинк. Так появился один из важнейших законов экологии, закон минимума Либиха: рост растений зависит от того элемента питания, который присутствует в минимальном количестве.
Позднее закон минимума был расширен на все живые организмы и все факторы. В современной трактовке этот закон звучит следующим образом: выносливость организма определяется самым слабым звеном в цепи его экологических потребностей. То есть жизненные возможности организма лимитируются экологическими факторами, количество и качество которых близко к необходимому для данного организма минимуму. Дальнейшее снижение этих факторов ведет к гибели организма.
Однако не только снижение интенсивности действия какого-либо фактора, но и превышение сверх допустимых пределов может оказывать лимитирующее воздействие на организмы. Например, эффективность фотосинтеза, как известно, определяется количеством света, падающего на поверхность зеленого листа. Излишнее затенение может привести к подавлению жизнедеятельности растения и даже к его гибели. Это способствует возникновению теневыносливых видов растений. Однако, если увеличить поток света, например для получения большего урожая теневыносливых растений, то эффективность фотосинтеза у них падает. Оптимальные пределы величины светового потока имеют практически все растения.
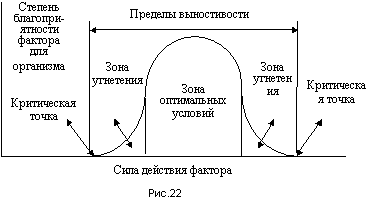
Поэтому вместо закона минимума в настоящее время чаще говорят о законе лимитирующих (ограничивающих) факторов: фактор, находящийся в недостатке или избытке, отрицательно влияет на организмы даже в случае оптимальных сочетаний других факторов. В соответствии с этим законом для каждого фактора по отношению к конкретному виду организмов можно вычертить диаграмму степени благоприятности данного фактора на организмы в зависимости от силы действия этого фактора. Почти всегда эта диаграмма имеет вид перевернутого колокола (рис.22). На ней можно отметить критические точки, определяющие пределы выносливости (толерантности, от латинского слова толеранция - терпение) данного вида организмов по отношению к рассматриваемому фактору (поэтому данная диаграмма называется кривой толерантности). Выход за эти пределы ведет к массовой гибели организмов. Зона вблизи максимума кривой называется зоной оптимальных условий. К ней обычно приурочена максимальная плотность популяции. За пределами этой зоны между критическими точками находятся зоны угнетения, в которых организмы находятся в состоянии стресса.
Таких кривых для данного вида организмов можно начертить множество - по каждому из факторов среды. Одна и та же среда обитания может предоставлять организмам такие сочетания различных факторов, что одни из них будут соответствовать зоне оптимальных условий для данного вида организмов, другие будут выходить за пределы этой зоны. Наиболее полно потенциальные возможности данного вида будут проявляться в том случае, если все факторы лежат в зоне оптимума. В соответствии с законом лимитирующих факторов выход за пределы толерантности хотя бы по одному из этих факторов чреват гибелью организмов. Точно также если один из факторов попадает в зону угнетения, то это будет иметь более существенное значение для организма, чем то, что все остальные факторы будут находиться в зоне оптимальных условий.
Правда, это правило имеет одно существенное ограничение, связанное с взаимодействием различных факторов. Так в 1930 г. Э.Рюбель сформулировал закон компенсации факторов: отсутствие или недостаток некоторых факторов может быть компенсирован другим близким фактором. В более общей формулировке он звучит следующим образом: одни факторы могут усиливать или смягчать действие других факторов. Например, недостаток света может в какой-то мере смягчаться повышенным содержанием углекислого газа в воздухе.
Именно лимитирующие факторы определяют обычно границы распространения видов (популяций), то есть их ареалы, от них зависит продуктивность сообществ, численность особей и многие другие параметры жизни.
Кривые толерантности для разных факторов могут иметь более широкую или более узкую форму, соответственно различаются и пределы толерантности организма по отношению к разным факторам. Организмы с широким диапазоном толерантности по отношению к данному фактору называются эврибионтами (от греческих слов эури - широкий и биос - жизнь). Организмы с узким диапазоном толерантности по отношению к данному фактору называются стенобионтами (от греческих слов стенос - узкий). Один и тот же организм может быть стенобионтом по отношению к одним факторам и эврибионтом по отношению к другим. Более того в разные периоды жизни одного организма его требования к среде могут существенно меняться. Например, по отношению к температуре личинки насекомых обычно стенобионтны, в то время как куколки и взрослые особи могут относиться к эврибионтам.
Эврибионты хорошо выдерживают широкий диапазон колебаний факторов среды, например, типичным эврибионтом является верблюд, способный жить не только в условиях пустыни, выдерживая значительные колебания температуры, недостаток влаги и пищи, но и в условиях умеренного пояса. Виды с широкими диапазонами толерантности обычно первыми заселяют новые районы, в которых произошли какие-то специфические изменения окружающей среды, для которых у природы пока еще не выработаны адаптации, то есть определенные приспособления к действию данных факторов. Если условия среды изменяются в малых диапазонах, то это способствует формированию у организмов четких адаптаций, иногда в ущерб ширине диапазона толерантности. При этом они оказываются способными нормально существовать в достаточно суровых по нашим меркам условиях, например, в полярных водах, где температура хотя и низкая (около 2оС), но достаточно стабильная, или в даже в жерлах вулканов. То есть эврибионты достойно выдерживают конкуренцию при достаточно широких и непредсказуемых колебаниях факторов среды. В более стабильных условиях в конкурентной борьбе как правило побеждают стенобионты.
Таким образом, появление в природе узких пределов толерантности можно рассматривать как форму специализации, в результате которой большая эффективность достигается в ущерб адаптивности организмов. Обычно с ростом стенобионтов в сообществе увеличивается разнообразие видов. То есть это есть одна из разновидностей дифференциации в природе.
Эта закономерность характерна для любого эволюционного процесса. В упоминавшемся уже процессе эволюции науки также можно наблюдать постепенную дифференциацию отдельных отраслей знания, которые все более сужают свой предмет. Специалисты в этих отраслях знания как правило характеризуются довольно узким кругозором, зато становятся отличными знатоками своего предмета. Подобные тенденции являются прямым следствием принципа оптимальности, так как в итоге система стремится к энергетически более выгодному состоянию. Узкая специализация способствует более эффективному преобразованию энергии, но при этом предъявляются повышенные требования к форме энергии на входе данной специализированной подсистемы. Такую форму должны обеспечить другие узкоспециализированные подсистемы, стоящие на предыдущих (а иногда и на более высоких) уровнях в цепи преобразования энергии. Но они в свою очередь по своему также требовательны к форме входной энергии. Так строится довольно сложная иерархичная, но достаточно запутанная, структура преобразования энергии данной системой, аналогичная трофической сети экосистем. Ее отличительная особенность в строгой увязке всех подсистем, в их взаимозависимости и в полной невозможности самостоятельного существования вне системы. Это как раз то, что отличает целостный живой организм от совокупности слабосвязанных друг с другом элементов. Именно благодаря узкой и взаимозависимой специализации отдельных особей можно считать живыми организмами такие биосистемы, как муравейник, термитник, улей, а также более крупные образования: коралловый риф, тропический лес, человеческий социум, биосфера в целом.
Дата: 2018-11-18, просмотров: 949.