Со структурой экологических ниш данной экосистемы тесно связана ее видовая структура, под которой понимается количество видов, образующих экосистему и соотношение их численностей.
Виды, преобладающие по численности особей, имеющие наибольшую значимость для экосистемы, называются доминирующими видами, или доминантами (от латинского слова доминантис - господствующий). Наряду с доминантами в экосистемах выделяют виды-эдификаторы (от латинского слова эдификатор - строитель), которые являются основными строителями среды. Обычно доминирующий вид является одновременно и эдификатором, в наших лесах это обычно ель (еловый лес), сосна (сосновый лес), береза (березовый лес) и т.п. Наряду с многочисленностью деревьев этих видов в соответствующих лесах, они еще определенным образом организуют среду для существования других организмов. Так в еловых лесах почва сильно затемнена и обладает повышенной кислотностью, это во многом определяет видовую структуру елового леса. В то же время под пологом леса может доминировать черника или кислица, которые не являются существенными эдификаторами, но хорошо характеризуют особенности условий в данной экосистеме, в частности степень увлажнения, плодородие почвы и т.д. Поэтому данные виды называют еще индикаторами. По растениям-эдификаторам и растениям-индикаторам называют биогеоценозы, например, ельник-кисличник (кислица указывает на оптимальное увлажнение и богатые почвы), ельник-черничник (менее богатые почвы и несколько избыточное увлажнение), ельник сфагновый (избыточное увлажнение, недостаток минеральных веществ и кислорода в почве) и т.д.
Важнейшим показателем видовой структуры экосистемы является видовое разнообразие. От видового разнообразия зависит, например, насколько успешно будут заполнены все экологические ниши данной экосистемы. Однако количество экологических ниш, как правило, гораздо больше, чем количество видов, населяющих данную экосистему. Как уже отмечалось, многие виды на разных стадиях жизненного цикла занимают разные экологические ниши. Поэтому для экологического разнообразия важно не столько количество видов, сколько видовое богатство, в которое помимо количества видов целесообразно включить еще и количество разных жизненных фаз внутри каждого вида. Кроме этого, нужно учесть еще и генетическое разнообразие, то есть генетическую изменчивость видов, без которой они оказались бы неспособными адаптироваться к изменяющейся среде. Большое значение для заполнения экологических ниш имеет и “поведенческое разнообразие”, зависящее от сложности организации организмов, их способности формировать в природе разного рода структуры (строить гнезда, объединяться в стаи, формировать симбиозы и т.п.), гибкости их инстинктов и уровня их психического развития (для высокоорганизованных животных), от наличия разума (достаточно вспомнить, что человек заполняет собой все большее количество экологических ниш). Все вместе это можно обозначить одним термином - экологическое разнообразие.
Если не принимать в расчет деятельность человека, то можно отметить, что экологическое разнообразие повышает устойчивость экосистем к воздействию неблагоприятных факторов среды. Например, видовое разнообразие обеспечивает дублирование разных функций. Так вид, представленный в данной экосистеме единицами особей, способен в случае гибели широко представленных видов резко увеличить свою численность и занять освободившиеся экологические ниши, сохранив жизнь экосистеме в целом. Этому значительно способствует генетическое разнообразие.
Деятельность человека также способствовала бы устойчивости экосистем, если бы он не возгордился, и не противопоставил себя природе. Присутствие разума дало бы экосистеме защиту даже от таких фатальных разрушителей, как верховые пожары, иногда полностью уничтожающие экосистемы, сопровождая это уничтожение сильной эрозией почвы. Человек, по-видимому, призван был быть мудрым правителем, потому-то природа, наделив его огромной силой, не сделала его хищником. Человек, в принципе, может жить в гармонии с природой, практически ничего от нее не требуя. Об этом свидетельствует опыт подвижников самых разных религиозных направлений. И вовсе не “золотой миллиард” способна прокормить Земля, а гораздо больше. Только не людей с такими запросами, как наши. Видно что-то не получилось в замысле природы. Или мы пока еще не выросли, и природа, как терпеливая мать, отдает нам все, приносит себя в жертву, в надежде на то, что когда мы созреем, отточим свой ум на собственных ошибках, тогда с избытком вернем ей свою благодарность. Ведь самое главное наше достояние - это способность отдавать тепло своих сердец тем, кто в этом нуждается. И тогда мы не только восстановим, но и приумножим жизнь на планете.
Пока же мы, наоборот, обедняем видовое разнообразие природы. Это всегда приносило человеку кратковременную выгоду. Например, наши предки выжигали целые леса, оставляя вместо древней экосистемы с богатым разнообразием голую равнину, богатую минеральными веществами, на которой ускоренными темпами начинает зарождаться новая экосистема. Но человек направлял этот процесс по-своему, засевая только семена конкретной монокультуры. Природа пыталась увеличить разнообразие, и вместе с данной монокультурой на свет появлялось разнотравье, но человек всячески боролся с этими “сорняками”, поддерживая бедноту разнообразия, потому что это позволяло получить гораздо большие урожаи.
Подобные антропогенные воздействия на природу, сопровождающиеся снижением разнообразия видов, особенно сильны в наше время. Помимо непосредственного отстрела животных, промысла рыб, рубки деревьев, каждый год из природы “изымается” огромное количество площадей под дороги, города, рудники и т.п. к этому можно добавить и то, что многие земли после «интенсивного сельского хозяйства» становятся пустынями. Все это способствует уменьшению разнообразия, так что количество видов растений и животных неуклонно сокращается прогрессирующими темпами.
В природе разнообразие видов накапливается за тысячи и даже миллионы лет. Огромную роль в видообразовании играют экологические ниши. Замечено, что видовое разнообразие в данной экосистеме зависит от ее возраста. По мере развития экосистемы видовое разнообразие сначала увеличивается, но после достижения зрелости системы оно стабилизируется или даже уменьшается. Однако подобная динамика характерна в основном для северных лесов и учитывает лишь относительно короткие периоды времени (в пределах 100-200 лет). Девственные районы северных лесов, например сибирской тайги, также не показательны, потому что суровые климатические условия значительно сдерживают экспансию жизни. Наиболее богаты видами тропические районы планеты. Именно здесь, в наиболее благоприятных для жизни и относительно стабильных условиях, сформировалось наибольшее количество видов планеты. Достаточно сказать, что практически все высшие животные нашей средней полосы имеют африканское происхождение.
Как уже отмечалось, каждая отдельная особь в экосистеме находится под двойным давлением: межвидовая конкуренция способствует сужению экологической ниши, а внутривидовая конкуренция давит в обратном направлении, заставляя данный вид расширять свое жизненное пространство. Так как все экологические ниши уже заняты, то единственный выход - “пойти туда, не знаю куда, и найти то, не знаю что”. Другими словами, для снятия противоречия нужно создать новую экологическую нишу с заранее неизвестными параметрами, например найти новый источник пищи. Но его не существует, так как иначе это говорило бы о наличии свободной экологической ниши, чего в девственной природе практически никогда не бывает. На помощь приходит механизм генетической изменчивости видов. Например, спасаясь от давления консументов, автотрофы вынуждены “изобретать” новые способы защиты. Когда это удается, то прессинг со стороны консументов ослабевает, и “новорожденный” вид приобретает некоторое преимущество перед близкородственными видами, поэтому он несколько теснит конкурентов, высвобождая для себя жизненное пространство. Иногда удается отвоевать достаточно много, может быть, даже поставив экосистему на грань потери устойчивости. Однако механизмы изменчивости консументов рано или поздно “нащупают” слабое место нового вида и дадут начало соответствующему виду консументов, вернув в прежнее русло равновесие в среде автотрофов. Все эти изменения постепенно пройдут по всей пищевой цепи, давая начало целой группе новых видов.
Конечно, это только один из возможных примеров. Аналогичные изменения могут возникнуть, например, при внедрении в экосистему какого-то вида из других экосистем, решающего свои внутривидовые проблемы путем миграции, если ему удается потеснить “родные” виды данной экосистемы. К подобным последствиям приводит и деятельность человека, типичным примером является история с кроликами, завезенными в Австралию. Правда, здесь роль хищника взял на себя человек, не дожидаясь, пока природа сама расставит все по местам. Многие “взрывы” видообразования приурочены к всевозможным потрясениям, обрушивающимся на экосистему: ураганы, извержения вулканов, падение метеоритов и т.п. При этом, если экосистема не потеряла своей устойчивости и не включились механизмы деградации, ввиду массовой гибели организмов может освободиться достаточно большое количество экологических ниш, что может дать начало формированию целой мутовки новых видов. Так или иначе, но чем старше экосистема, расположенная в благоприятных для жизни условиях, тем больше видов она успевает сформировать.
Рассмотренные примеры создают впечатление, что видообразование инициируется “снизу”, то есть со стороны конкретных особей. Это типично редукционистский подход. С точки зрения холизма инициатором видообразования является экосистема. Именно она в своем эволюционном развитии закрепляет или отсеивает всевозможные мутации, запуская в действие механизмы видообразования. В свою очередь, ее динамика направляется, как правило, со стороны надсистем более высокого порядка. Так самые мощные и коренные перестройки видовых структур экосистем биосферы вызваны климатическими изменениями на планете.
Способность природы увеличивать разнообразие может явиться мощным оружием против человека. Деятельность человека по отношению к природе можно сравнить с потрясением, грозящим погубить жизнь. Такое в истории планеты случалось не однажды, о чем свидетельствуют бедные жизнью прослойки между геологическими слоями, когда жизнь, похоже, на время замирала, а потом взрывоподобно возрождалась, но уже в новых формах. То есть адаптации к подобным потрясениям жизнь уже сформировала. В соответствии с принципом Ле Шателье-Брауна, жизнь всегда пытается компенсировать неблагоприятные воздействия на нее.
Рассмотрим один из механизмов, с помощью которых природа должным образом реагирует на подобные потрясения. Чем интенсивней человек обедняет разнообразие, тем большее количество экологических ниш становится свободными. При этом уменьшается и межвидовая конкуренция, и оставшиеся виды одного трофического уровня замещают исчезнувшие, заполняя освободившиеся ниши. При этом, как правило, действует схема: мелкий сменяет крупного, эволюционно менее организованный сменяет высокоорганизованного, генетически изменчивый сменяет менее изменчивого. Например, копытных в степи могут сменять грызуны, а в некоторых случаях - насекомые (вспомните нашествия саранчи). Исчезающих хищников может заменить крыса.
Особенно ярко этот механизм заметен на популяциях микроорганизмов, возбудителей человеческих болезней. Период деления бактерий, например, может составлять всего несколько часов. У вирусов динамика еще более высокая. Это значит, что видообразование в этой среде может происходить буквально на глазах. Успехи медицины по уничтожению возбудителей болезней свидетельствуют, что на смену “побежденному” виду микроорганизмов, через достаточно короткое время приходит другой, уже адаптированный к действиям препаратов, вызвавших гибель его предшественника.
Таким образом, фармакология никогда не стоит на месте, она вынуждена постоянно совершенствовать медицинские препараты, которые приносят временные победы, но неизбежно стимулируют эволюционный прогресс в мире возбудителей болезней. Причем период времени от появления нового лекарства до соответствующей мутации в мире микроорганизмов все более сокращается и грозит свести на нет саму целесообразность существования фармакологии. Это вызвано тем, что освободившиеся экологические ниши занимают зачастую более примитивные микроорганизмы, чем их предшественники.
Так после изобретения антибиотиков главным врагом человека стали не простейшие грибки и одноклеточные, а вирусы. Есть первые симптомы того, что на смену вирусам грядут ретровирусы - довирусные, более древние формы жизни, строящие свою организацию не на основе молекулы ДНК, а на основе РНК. С ними бороться сложнее. Одним из наиболее известных представителей этой формы жизни является вирус (более правильно ретровирус) СПИДа. За 13 лет до его открытия была предсказана вероятность “гриппоподобного заболевания с высокой летальностью”.
Интересно, что анализ путей распространения СПИДа в Африке совпадает с бывшими путями распространения оспы. Но оспу мы победили. Это не значит, что человек избавился от возбудителей болезней. Все экологические ниши должны быть заняты. И если человек с этим не хочет мириться, единственное что он может сотворить - это мощную эволюцию, взрыв разнообразия новых возбудителей болезней, с помощью которых рано или поздно природа установит равновесие в биосфере, нарушенное человеком.
Глава 12.
Динамика экосистем
Цель: особенности жизненного цикла надсистем
Несмотря на то, что многие экологи не склонны относить экосистемы к “сверхорганизмам”, между развитием экосистем и развитием живых организмов имеется множество параллелей. Как и любые организмы, экосистемы рождаются, старятся и умирают, точнее, структурно перестраиваются или замещаются другими экосистемами. Выявлена масса закономерностей протекания этих процессов, которые своей повторяемостью свидетельствуют о наличии достаточно однозначных “программ” развития экосистем в том или ином регионе.
На каких носителях “записаны” эти программы? Что является генетическим кодом онтогенеза (индивидуальное развитие) экосистем? Как уже не раз отмечалось, ход развития определяется совокупностью факторов среды. В одинаковых условиях развиваются одинаковые по структуре экосистемы. Это своего рода давление надсистемы на эволюционирующую экосистему (макроуровень). Кроме того, следует учесть и характер законов природы, “зашитых” в формах организации материи, из которых строится экосистема, лежащих в основе всех механизмов природных явлений (микроуровень). Кто-то склонен переоценивать роль надсистемы в эволюции подсистем, считая, что развитие подсистем целиком подчинено требованиям более высокоуровневой организации (холистический подход). Кто-то, наоборот, категорически отрицает роль каких-либо надсистем, сводя всю динамику экосистем к механизмам межвидовых и внутривидовых отношений, генетической изменчивости, к борьбе за выживание и т.п. (редукционистский подход). Ни тот ни другой путь не дает полного понимания происходящих процессов, о чем свидетельствует бесконечный спор между сторонниками обеих концепций (аналогично спору между идеалистами и материалистами в философии).
К счастью, современная наука, похоже, открыла причину своих неудач в понимании мира. Принцип дополнительности находит признание и применение во все более широких областях знания. Только в единстве холизма и редукционизма возможно понимание сути явлений. Каждый подход по-своему прав, но только в их единстве возможно достижение полноты истины.
Виды динамики экосистем
Любая экосистема постоянно находится в состоянии динамики (изменений), вызванной как изменением факторов среды, то есть внешними возмущениями, так и внутренними процессами. При этом удобно различать циклическую динамику и поступательную. К разряду циклической динамики можно отнести:
1) суточную динамику, связанную с изменением активности растений и животных в течение суток: изменения в фотосинтезе, транспирации (испарение воды с поверхности листьев), у животных это смена периодов активности и сна и т.п.;
2) сезонную динамику (смена периодов вегетации и покоя), связанную с сезонными колебаниями: смена времен года, периодов дождей и засухи, муссонная периодичность и т.п.;
3) многолетнюю цикличность, вызванную, например, периодичностью солнечной активности (период порядка 10-11 лет), или же какими-то другими колебаниями с многолетним периодом, например, цикл Эль-Ниньо, который приводит к усилению ураганов, обрушивающихся на Южную Америку с периодичностью 5-7 лет, и т.п.
Особого внимания заслуживает поступательная динамика экосистем, или развитие. Обычно развитие сопровождается последовательным рядом изменений видовой и трофической структур экосистемы, всей ее организации, что позволяет даже говорить о последовательной смене экосистем, поэтому данный процесс называется сукцессией (от латинского слова сукцессио - преемственность, наследование). В зависимости от причин сукцессии различают экзодинамические (от греческого слова эксо - снаружи) сукцессии, вызванные внешними по отношению к данной экосистеме факторами, и эндодинамические (от греческого слова эндон - внутри) сукцессии, вызванные внутренними механизмами экосистемы (в теории управления различают вынужденную и свободную составляющие динамического процесса, которые соответствуют двум видам сукцессии экосистем).
Экзодинамические сукцессии могут быть вызваны изменениями климата, понижением уровня грунтовых вод, подъемом уровня мирового океана и т.п. Такие смены могут длиться столетиями и тысячелетиями. Они связаны в основном с действием механизмов адаптации экосистемы к факторам внешней среды, которые в свою очередь базируются на механизмах адаптации живых организмов экосистемы.
Эндодинамические сукцессии приводятся в действие особыми законами, механизмы которых до сих пор во многом непонятны. Известно, что на любом, даже абсолютно безжизненном, субстрате типа песчаных дюн или затвердевшей лавы, рано или поздно расцветает жизнь. При этом формы жизни, точнее, типы сообществ, в данном пространстве последовательно сменяют друг друга, постепенно усложняясь и увеличивая видовое разнообразие, формируя так называемый сукцессионный ряд, состоящий из последовательных стадий, отмечающих смену одного сообщества другим. Сукцессионный ряд заканчивается стадией зрелости, на которой экосистема изменяется очень мало. Экосистемы на этой стадии называются климаксными (от греческого слова климакс - лестница).
|
| Рис.23 |
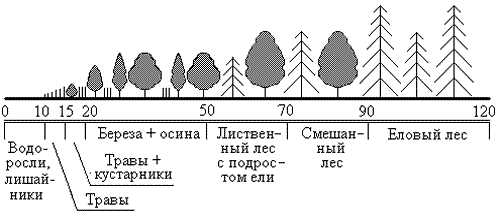
Продолжительность сукцессии от зарождения экосистемы до стадии климакса может составлять до сотен и даже до тысяч лет. Столь длительная продолжительность связана в основном с необходимостью накопления питательного вещества в субстрате. Изначально на безжизненном субстрате поселяются так называемые пионерные виды, типа лишайников и корковых водорослей (рис.23). За 5-10 лет они несколько обогащают субстрат питательными веществами, формируя зачатки почвы. Затем на этих еще совсем бедных почвах поселяются травы, еще более обогащая почву. Лет через 15 от начала сукцессии на когда-то безжизненном пространстве поселяются первые кустарники, которые постепенно вытесняются лиственными светолюбивыми деревьями, чаще всего березой и осиной, которые характеризуются быстрым ростом. К 50-летнему возрасту в молодом лиственном лесу выделяются наиболее сильные деревья, которые затеняют более слабую поросль, которая погибает, давая возможность поселиться под пологом лиственного леса поросли ели. Ель более теневынослива, под защитой лиственных деревьев она постепенно догоняет их в росте, отвоевывая у них жизненное пространство. Где-то к 70 годам экосистема достигает стадии смешанного елово-лиственного леса. Лиственные деревья к тому времени успевают состариться, и постепенно ель выходит на первый ярус, затеняя и изреживая всю лиственную растительность. К 90 годам данная экосистема достигает стадии климакса, для которой характерно практически полное отсутствие лиственных деревьев, ель становится доминирующим видом-эдификатором, формируя особым образом всю жизнь населяющего данную экосистему сообщества.
Это типичный пример сукцессии, характерный для южной тайги. Как ни странно, в одном и том же регионе практически все сукцессии развиваются по довольно похожим сценариям и заканчиваются в итоге одним и тем же климаксом. Конечно, в зависимости от конкретных особенностей среды возможны некоторые вариации общего сценария, но они не принципиальны. Таким образом, каждый достаточно крупный регион характеризуется своей программой развития экосистем и своим особым климаксом, который в данном смысле называется моноклимаксом. Это еще один аргумент в пользу биополевого механизма координации жизни в биосфере - каждый регион планеты развивается в соответствии с характерной для него программой и имеет свою характерную структуру, призванную обеспечить определенную функцию, закрепленную за данным регионом в едином организме биосферы (или планеты).
Правда, справедливости ради надо сказать, что у автора концепции моноклимакса, американского эколога Клементса, имеется достаточное количество оппонентов. Может быть, это связано с тем, что любая крайне полярная точка зрения всегда оказывается односторонней. Концепция моноклимакса хорошо укладывается в холистический подход, однако сторонники исследования детальных механизмов природных явлений приводят множество примеров, когда в районах с известным моноклимаксом формируются совершенно другие климаксные экосистемы. Например, в южной тайге наряду с еловыми лесами могут формироваться луговые и сосновые климаксные экосистемы. Сторонники моноклимакса склонны рассматривать их как промежуточные стадии, которые рано или поздно обязательно должны прийти к моноклимаксу, если для этого не будет таких препятствий, как, например, вмешательство человека. Так луговые экосистемы, похоже, существуют только за счет регулярного скашивания молодой древесной поросли. Сосновые леса могут возникать и без участия человека, особенно на крайне бедных почвах, типа песчаных и щебнистых почв, куда более требовательная к почвам ель внедриться не может. Однако по мере накопления плодородия почв, ель, как более сильный эдификатор, скорее всего, вытеснит сосну и создаст моноклимаксную экосистему.
Проверить это достаточно сложно, так как подобного рода сукцессии растянуты на тысячи лет. Иногда экологам улыбается удача и они обнаруживают экосистемы, где можно непосредственно “наблюдать” такие тысячелетние сукцессии. Это позволяет выявить некоторые характерные особенности сукцессионного процесса. Так например, замечательной природной лабораторией являются дюны у южной оконечности озера Мичиган (США, штат Индиана). Озеро постепенно отступает, оставляя после себя все более молодые песчаные дюны, абсолютно безжизненные, которые постепенно “осваиваются” жизнью. Процесс этот длится уже тысячи лет, поэтому разноудаленные от кромки воды дюны находятся на разных стадиях сукцессии, что позволяет проследить весь ход сукцессионного процесса. Первые поселенцы на дюнах - это прибрежные злаки, ива, песчаная вишня, хлопковое дерево и животные: пауки, кузнечики и т.п. За ними следуют сухие сосновые леса, затем сухие леса из черного дуба, и, наконец, на самых старых дюнах - влажные леса из дуба, бука и клена, для которых характерна мощная богатая гумусом почва, населенная дождевыми червями и улитками. Для достижения климаксного сообщества на дюнах требуется около 1000 лет, что примерно в пять раз больше, чем это требуется при более лучших, чем дюны, исходных условиях.
В зависимости от исходных условий принято различать первичные сукцессии, которые начинаются на абсолютно безжизненных субстратах, например, на дюнах, и вторичные сукцессии, начинающиеся с более благоприятных стартовых условий, например, после пожара, вырубки леса или на заброшенном поле. Вторичные сукцессии могут начинаться с более поздних пионерных видов, например, с трав или кустарников. Так например, сукцессии на заброшенных полях начинаются с однолетних трав, бывших ранее сорняками. Затем они сменяются многолетними травами, кустарниками и деревьями. Ввиду большего исходного плодородия почвы такие сукцессии достигают климакса гораздо быстрее, чем в случаях первичной сукцессии. В последнее время для биосферы характерны именно вторичные сукцессии, что связано главным образом с деятельностью человека.
Вмешательство человека в природу ведет зачастую также к явлениям дигрессии (от латинского слова дигрессион – отклонение), когда климакс экосистемы (устойчивое равновесное состояние) достигается на более ранних стадиях сукцессии, вследствие чего экосистемы значительно упрощаются. В отличие от моноклимакса эта стадия называется катаценозом (от греческого слова ката – вниз, против). Типичным примером является пастбищная дигрессия, когда вместо леса, на данном пространстве формируется обширная луговая экосистема. Чем дальше отступает пояс лесов, тем реже достигают данного пространства семена кустарников и деревьев. Регулярный выпас скота не дает развиться молодым деревьям. Постепенно на этом месте вдали от лесов формируется особая зона, богатая гумусом, где никогда (точнее, очень долго) не вырастет лес, даже если человек оставит этот район. Так, вероятно, формировались степи, на которых древние кочевые племена пасли свои табуны и стада. Попытки возобновить леса в степной зоне требуют немалых усилий, так как помимо колонии деревьев, лес характеризуется особой структурой симбиотических связей, особенно между конями деревьев и грибницей (микроза). Поэтому лес может постепенно “наступать” на степь. Но, чтобы лес самопроизвольно возник посреди степи - случай практически невозможный. Для того, чтобы лес заменил степи после ухода человека потребуются, вероятно, десятки тысяч лет.
Следует различать автотрофные и гетеротрофные сукцессии. Все описанные выше примеры относятся к автотрофным сукцессиям, так как они протекают в экосистемах, где центральным звеном является растительность (фитоценоз). Динамика гетеротрофов целиком подчинена динамике автотрофов - смена животных сообществ зависит от смены растительных сообществ. Автотрофные сукцессии могут длиться теоретически вечно, так как они постоянно подпитываются энергией Солнца.
В гетеротрофных сукцессиях участвуют только животные (гетеротрофы, консументы). В этот процесс могут быть также вовлечены и мертвые растения, например, поваленные деревья, пни и т.п., которые являются, как правило, источником энергии для гетеротрофной сукцессии. Примером такой сукцессии может служить разрушение поваленного дерева. Первыми на мертвом или ослабленном дереве поселяются насекомые-короеды, которые повреждают кору, давая возможность проникнуть в места повреждений спорам грибов. Грибы-пионеры размягчают древесину, окрашивая ее в разные цвета. Это облегчает появление следующей волны насекомых, которые питаются уже древесиной (ксилофаги). В поврежденной ими древесине развиваются грибы-деструкторы, способствующие появлению мягкой гнили. Затем их сменяют грибы-гумификаторы, превращающие гнилую древесину в гумус. На всех стадиях гетеротрофной сукцессии также идет соответствующая смена видов микроорганизмов. В конечном итоге все органическое вещество разлагается до конечных продуктов: минеральных веществ и углекислого газа. Аналогичные процессы идут и при разложении трупов животных, а также в экскрементах животных, в лесной подстилке, в загрязненных водах и т.д.
|
| Рис.24 |
Гетеротрофная сукцессия предполагает обязательное наличие определенного запаса энергии, аккумулированной в органическом веществе. Она заканчивается вместе с исчерпанием ресурса энергии, то есть после полного разложения исходного субстрата. После этого экосистема перестает существовать. То есть для нее не определено понятие климакса. В отличие от биогеоценозов, такие экосистемы смертны.
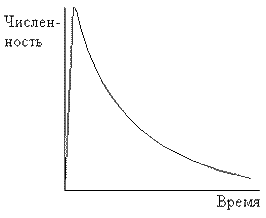
Гетеротрофная сукцессия хорошо ассоциируется с обществом, зависящим от ископаемого горючего. Динамика гетеротрофной сукцессии описывается кривой с быстрым ростом числа организмов, пока не будет достигнут некоторый максимум, затем количество организмов постепенно уменьшается по мере исчерпания энергетического ресурса (рис.24). Какого-то устойчивого состояния (климакса) добиться не удается. Такое общество быстро прогрессирует, но тем не менее оно изначально обречено на вымирание. Мы уже “сняли сливки” с большинства месторождений. Дальнейшая их эксплуатация со временем будет требовать все больших вложений энергии. Поэтому эффективность добычи полезных ископаемых будет неуклонно падать. Вместе с этим будет падать и жизнеспособность цивилизации, построенной на гетеротрофной сукцессии, если, конечно, еще раньше не произойдет катастрофических изменений, например, массового психоза - людям свойственно искать виновников своих проблем. Поэтому мы огромное количество усилий направляем на поиск новых источников энергии. Но даже если мы научимся управлять термоядерным синтезом, это не изменит нашей деструктивной сущности. Мы только несколько отодвинем в будущее развязку, как это мы делали уже не раз, но стабилизировать гетеротрофную сукцессию нам не удастся. Это даже если не учитывать, что в случае успеха в области энергетики перегрев биосферы неизбежен.
Дата: 2018-11-18, просмотров: 954.