Понятие энергии определяется как способность совершать работу. Хотя вся современная наука проникнута этим понятием, природа энергии до сих пор до конца не понята. Особенно ярко это проявилось после того, как мир узнал, что E = mc2.
Впервые наиболее полно понятие энергии было проработано в термодинамике, что вылилось в формулировку двух наиболее основополагающих законов, описывающих свойства энергии:
1) первый закон термодинамики (принцип сохранения энергии) - энергия может переходить из одной формы в другую, но она никогда не исчезает и не создается заново;
2) второй закон термодинамики (принцип роста энтропии) - все реальные процессы превращения энергии сопровождаются ростом энтропии, то есть переходом энергии в более рассеянное состояние.
Энтропия, или дословно способность к превращению, есть величина еще более непонятная, чем энергия. Некоторые вопросы, касающиеся понятия энтропии, мы уже рассмотрели в предыдущих главах. Здесь мы несколько дополним его, акцентировав внимание именно на энергетической стороне вопроса. Для этого рассмотрим сначала, каким образом термин энтропия вошел в научный лексикон.
Известно, что во всех естественных процессах теплообмена тепло передается только от тела с большей температурой к телу с меньшей температурой и никогда наоборот. То есть если от тела 1 с температурой Т1 и энтропией S1=Q1/Т1 отводится к телу 2 с температурой Т2 (Т1 > Т2) и энтропией S2 = Q2/Т2 некоторое количество теплоты dQ, достаточно малое, чтобы температуры обоих тел не уменьшились, то энтропия тела 1 изменится (уменьшится) на величину dS1 = dQ/Т1 (здесь dQ < 0, следовательно, dS1 < 0), а энтропия тела 2 изменится (увеличится) на величину dS2 = dQ/Т2 (здесь dQ>0, следовательно, dS2 > 0), причем так как Т1 > Т2, то по абсолютной величине |dS1| < |dS2|, поэтому общая энтропия двух тел S = S1 + dS1 + S2 + dS2 = S1 - |dS1| + S2 + |dS2| > S1 + S2, то есть в процессе теплопередачи суммарная энтропия двух тел возрастает и никогда не убывает. Это и есть формулировка второго закона термодинамики. То есть энтропия - это величина, характеризующая направление естественных процессов теплопередачи и, как выяснилось, вообще любых процессов преобразования энергии.
Энтропию называют тенью энергии. Как мы уже говорили, в более широком смысле под энтропией понимают меру качества, то есть меру концентрации и упорядочения энергии. Так тепловая энергия с большей температурой обладает меньшей энтропией S = Q/T, то есть большим качеством, чем такое же количество тепла при меньшей температуре. Поэтому по мере понижения температуры рабочего тела, например, пара, до температуры окружающей среды, можно попутно превратить часть тепловой энергии в механическую работу (тепловая машина). Чем больше качество энергии, то есть, чем больше превышение температуры пара над температурой окружающей среды, тем большее количество работы можно получить. Разные виды энергии обладают разным качеством. Так, например, упорядоченное движение частиц твердого тела (механическое движение) обладает большим качеством, чем хаотичное движение этих же частиц с той же средней скоростью (тепловое движение). Поэтому любое механическое движение при наличии трения сопровождается самопроизвольным превращением части механической энергии в тепловую. Поэтому иногда используют такие формулировки второго закона термодинамики: невозможно создать машину с КПД = 1; все естественные процессы идут в направлении ухудшения качества (деградации) энергии.
Когда мы говорим об энергии, особенно в контексте, связанном с энергетическим кризисом, следует помнить, что энергии на Земле вполне достаточно. Теплоход, идущий по океану, идет по морю энергии. Тем не менее он вынужден вести с собой запас угля, потому что энергия, запасенная в океане обладает низким качеством. Для полезного использования нужна именно высококачественная энергия, энтропия которой ниже энтропии энергии, рассеянной в окружающей среде. Энергию океана можно использовать только при наличии холодильника с более низкой температурой, чем температура океана. В качестве такого холодильника может выступать только космос, в который Земля “сбрасывает” излишки тепла. За счет этого ресурса во многом формируется энергия ветров.
Именно разность энтропий на входе и выходе энергетического потока порождает фактор, который мы обозначаем понятием силы, приводящей в движение все процессы в природе. По сути дела, любая сила имеет энтропийную природу. Рассмотрим пример с обычным футбольным мячом. Если он туго накачан, то можно выделить область, ограниченную поверхностью мяча, где концентрация частиц (носителей энергии) больше, чем в окружающей среде. Накачивая мяч, мы создаем неравновесие в системе, упорядочивая частицы в пространстве и увеличивая концентрацию энергии. По второму закону термодинамики система стремится к состоянию с максимумом энтропии. Это воспринимается в окружающем мире как проявление силы давления. Чем дальше система от равновесного состояния, тем больше сила. Эта сила создает поток энергии, направленный в сторону уменьшения неравновесия в системе. Остановить этот поток может только то, что при его появлении по каким-то причинам будет уменьшаться энтропия. Но растяжение резины поверхности мяча как раз и приводит к уменьшению энтропии. Понять это можно, рассмотрев строение молекулы каучука. Она представляет собой длинную полимерную цепь, свернутую в клубок случайным образом. Попытка выпрямить ее путем растяжения резины приводит к увеличению порядка в молекуле, то есть к уменьшению энтропии. Таким образом, два противоречивых фактора оказывают противоположные воздействия, в системе устанавливается такое состояние, которое соответствует локальному максимуму энтропии.
Наличие упорядоченных структур типа кристаллических решеток, живых организмов и т.п., способствует упорядочению движения частиц за счет уменьшения их степеней свободы. Поэтому принцип роста энтропии требует роста количества степеней свободы в каждом реальном процессе превращения энергии. Поэтому все упорядоченные структуры имеют тенденцию к разрушению. “Все разрушается, все умирает, все приходит в хаос” - это еще одна формулировка второго закона термодинамики.
Правда, помимо такого разрушения есть еще один способ увеличения количества степеней свободы - усложнение структуры системы. Именно по этому пути движется глобальный эволюционный процесс. При этом природа никогда не стремится достичь полного хаоса на данном уровне системной иерархии. В этом случае эволюция Вселенной остановилась бы достаточно быстро. Обычно в пределах данного иерархического уровня открываются некоторые устойчивые структуры, из которых строятся более высокие иерархические уровни, характеризующиеся большими значениями максимально возможной энтропии, чем на предыдущем уровне. Это дает возможность непрерывному росту энтропии.
Так обычно тенденция к возникновению хаоса реализуется в стремлении вещества к рассеянию (например, растворение сахара в воде). Но в случае сложных органических соединений больший хаос (рассеяние энергии) может быть достигнут именно при концентрации вещества. Например, капли масла, рассеянные в воде, стремятся слиться в одну большую каплю. Дело в том, что молекулы воды “окутывают” молекулу углеводорода своеобразной упорядоченной оболочкой. Поэтому чем больше поверхность масла, тем более упорядоченными оказываются молекулы воды, чего природа допустить не может. Поэтому в хаосе движения капель они обязательно рано или поздно примут состояние с наименьшей поверхностью, то есть сольются в одну большую каплю.
Именно это, вероятно, послужило в свое время началом одноклеточной жизни. Так в растворе белковых молекул формируются так называемые коацерватные капли, имеющие стабильную и иногда достаточно сложную структуру и поглощающие из раствора строго определенные вещества.
В биосистемах стремление к хаосу реализуется еще в более сложных механизмах. Примером тому может служить процесс деления клеток. Производство энтропии за счет протекания внутриклеточных процессов пропорционально объему клетки V, а отток энтропии из клетки пропорционален площади ее поверхности S. Если клетка имеет форму шара, то V = 4p  r3/3, S = 4 r2. Прирост энтропии в клетке
r3/3, S = 4 r2. Прирост энтропии в клетке  S = A
S = A  4p r3/3 - B 4 r2. При малых радиусах прирост энтропии S < 0. С ростом клетки ее радиус увеличивается, пока не достигнет некоторого критического значения при r = 3B/A, характеризующегося S = 0. В случае дальнейшего роста энтропия в клетке будет расти S > 0. Чтобы не допустить этого, она должна разделиться, иначе она погибнет от голода, перегрева и отравления своими же отходами.
4p r3/3 - B 4 r2. При малых радиусах прирост энтропии S < 0. С ростом клетки ее радиус увеличивается, пока не достигнет некоторого критического значения при r = 3B/A, характеризующегося S = 0. В случае дальнейшего роста энтропия в клетке будет расти S > 0. Чтобы не допустить этого, она должна разделиться, иначе она погибнет от голода, перегрева и отравления своими же отходами.
Существуют и другие механизмы, решающие данную проблему. Клетка может увеличить площадь своей поверхности, например, приобрести форму эллипсоида, цилиндра (палочки) или нити, образовать корнеподобные выросты, ложноножки и т.п. Многоклеточные организмы решают подобную проблему аналогичным образом. У растений увеличивается поверхность листьев и корней. У животных в отличие от растений подобное увеличение поверхности упрятано обычно внутрь организма, чтобы не мешать движению. Достаточно вспомнить развитые поверхности кишечника, органов дыхания, кровеносной системы и т.п. Например, общая поверхность всех эритроцитов взрослого человека составляет около 3000 м2, общая длина всех капилляров - около 100 000 км и т.д.
Нечто аналогичное происходит и в рамках таких сверхорганизмов, как экосистемы. Здесь дифференциация достигается путем увеличения экологических ниш и разнообразия видов, населяющих данную экосистему, удлинением и усложнением пищевых цепей, совершенствованием внутривидовых и межвидовых отношений и т.п. Все это есть следствие принципа роста энтропии.
Таким образом, разрушение структуры, требуемое принципом роста энтропии, является необходимым компонентом жизненного процесса. Но жизнь научилась использовать разрушение во благо, поэтому разрушение не обязательно сопровождается гибелью биосистем. “Умеренное разрушение”, на которое накладываются определенные запрограммированные ранее ограничения, приводит к расширению и усложнению жизни. Наиболее характерно в этом отношение деление клетки. Здесь смерть и рождение слились в одном процессе. Очень ярко об этом свойстве жизни выразился Ричард Бах: “Там, где глупец видит смерть гусеницы, мудрец видит рождение бабочки”.
Энергетика экосистем
Если движение вещества зачастую организуется в глобальный круговорот, захватывающий многие экосистемы биосферы, то движение энергии удобно рассматривать на примере какой-то одной экосистемы. Достаточно крупные экосистемы, такие как биогеоценозы, имеют все промежуточные уровни, которые проходит энергия при движении ее от состояния солнечного света до состояния тепла, которое сначала утилизируется в буферных зонах биосферы (атмосфера, гидросфера, литосфера), а затем излучается в космическое пространство (в инфракрасной части электромагнитного спектра).
Вывод энтропии из организма есть непременное условие его существования. Все процессы жизнедеятельности сопровождаются ростом внутренней энтропии организма Si > 0. Чтобы не погибнуть, клетка должна потребить из окружающей среды отрицательную энтропию (негэнтропию, информацию) Se < 0, что равносильно выводу энтропии из организма. Для этого обычно используется энергия химических реакций. Нужно взять из окружающей среды необходимые компоненты (пища) и создать условия для протекания реакции, продуктами которой должны стать вещества, содержащие в своей структуре больше энтропии, чем исходные компоненты. Обычно в этих реакциях разрушаются структуры более сложных молекул, например, молекул белка, жиров или углеводов. Затем эти продукты распада удаляются из организма. Себе же организм оставляет нечто, характеризующееся разницей энтропии исходных компонентов и энтропии продуктов реакции Se = Sисх-Sпрод < 0. Это нечто мы называем свободной энергией, которая по отношению к данному организму обладает отрицательной энтропией (негэнтропией), и за счет которой приводятся в движение внутренние упорядоченные процессы.
Например, глюкоза сгорает в организме, образуя двуокись углерода и воду. Это один из самых универсальных процессов, который лежит в основе дыхания и пищеварения. Разумеется, при сгорании глюкозы внутри организма не возникает пламени: живой организм устроен гораздо сложнее и тоньше, чем очаг, он извлекает свободную энергию химическими средствами, не допуская более знакомого нам расточительного сгорания. Продукты реакции, двуокись углерода и вода удаляются из организма при дыхании, потовыделении, экскрементации и т.п. Высвобожденная энергия претерпевает ряд превращений, обеспечивая тем самым протекание всех физиологических процессов, двигательных функций и т.п. Эту часть энергии рассматривают как траты на дыхание. Частично деградируя в каждом таком превращении, энергия постепенно полностью переходит в тепло, которое после этого удаляется из организма в окружающую среду.
Однако не вся свободная энергия, полученная в подобных экзотермических реакциях, проходит через организм подобным путем. Часть энергии используется на организацию ряда эндотермических реакций, то есть связывается в сложных молекулярных структурах. В первую очередь это реакции синтеза необходимых белков, нуклеиновых кислот и т.п. В данном случае эта доля свободной энергии идет на строительство и “ремонт” организма, то есть на упорядочение внутренней структуры. Эта энергия, накопленная в веществе организма, называется продукцией.
Некоторая доля пищи не усваивается организмом, следовательно, из нее не высвобождается энергия. Эта энергия выводится из организма вместе с экскрементами и впоследствии высвобождается из них уже другими организмами.
Ввиду наличия в своей структуре сложномолекулярных соединений, данный организм может служить пищей для другого организма. При этом его структура подвергается механическому и химическому разрушению. Высвободившаяся при этом свободная энергия используется так же, как в вышеописанном случае. Таким образом, формируется так называемая пищевая или трофическая (от греческого слова трофе - питание) цепь, в которой происходит перенос энергии через ряд организмов путем поедания одних организмов другими.
Трофическая цепь, как правило, иерархична, то есть состоит из последовательности уровней, называемых трофическими уровнями. Организмы, стоящие на каждом трофическом уровне, приспособлены природой для потребления определенного вида пищи, в качестве которой выступают организмы предыдущего трофического уровня (или нескольких предыдущих уровней). В принципе, организмы с более высоких трофических уровней также могут служить пищей на данном уровне, но это не является характерным, так как каждый следующий уровень трофической цепи аккумулирует в себе более качественную энергию и поэтому играет регулирующую функцию по отношению к нижним уровням, о чем будет подробнее сказано ниже. Другими словами, чем дальше трофический уровень от начала цепи, тем сильнее влияние организма на окружение, тем больше его возможности.
Трофические цепи можно разделить на два основных типа: пастбищную цепь и детритную цепь.
На вершине пастбищной цепи стоят зеленые растения (рис.19). Они не могут высвобождать энергию путем разрушения органики с предыдущего трофического уровня, поэтому единственным источником энергии является солнечный свет. При этом используются только достаточно энергичные фотоны длиной волны 380-710 нм, что близко к видимой части спектра (наиболее сильно поглощаются синий и красный цвета, зеленый свет поглощается слабее). Эту энергию называют фотоактивной радиацией (ФАР). В качестве строительного материала, то есть исходных компонентов для синтеза, используются простейшие минеральные и органические вещества, рассеянные в почве и в воздухе. К наиболее важным компонентам относится углекислый газ, являющийся продуктом жизнедеятельности всех организмов планеты. Именно здесь происходит возвращение в круговорот биологического углерода.
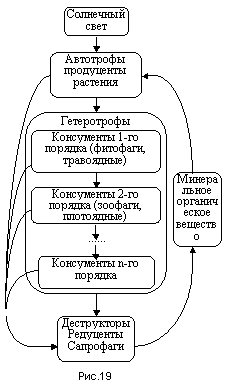
Так как зеленые растения “никого не едят” и все необходимое для их жизни синтезируют сами (конечно с участием солнечного света), их называют автотрофами (“самопитающимися”). Все остальные уровни трофической цепи существуют за счет энергии, накопленной в органике зеленых растений. Поэтому по отношению к трофической цепи растения называют продуцентами, то есть создающими первичную продукцию. Организмы на всех остальных уровнях трофической цепи называются консументами (потребителями) первого, второго и т.д. порядка в зависимости от трофического уровня. Так как эти организмы не могут сами синтезировать органику и вынуждены питаться другими организмами, их называют гетеротрофами (питающийся другими).
На втором уровне пастбищной цепи стоят обычно фитофаги, то есть животные, питающиеся растениями, в частности травоядные. Третий и более высокий уровни занимают хищники или зоофаги (питающиеся животными). Иногда эта цепочка может быть достаточно длинной, особенно в водоемах. Например, фитопланктон - зоопланктон - личинки насекомых - мелкая рыба - крупная рыба - хищные животные суши - животные, питающиеся падалью. Могут быть и промежуточные звенья.
Любая пастбищная цепь переходит в детритную цепь. Термин детрит означает «продукт распада» от латинского слова deterere - изнашиваться. Он позаимствован из геологии, где им называют продукты разрушения горных пород. В экологии детритом называют органическое вещество, вовлеченное в процесс разложения.
Уже уровень животных-падальщиков можно считать началом детритной цепи. В отличие от пастбищной цепи размеры организмов при движении вдоль детритной цепи не возрастают, а наоборот, уменьшаются. Так на втором уровне могут стоять насекомые-могильщики. Но самыми типичными представителями детритной цепи являются грибы и микроорганизмы, питающиеся мертвым веществом и довершающие продукт разложения биоорганики до состояния простейших минеральных и органических веществ, которые затем в растворенном виде потребляются корнями зеленых растений в вершине пастбищной цепи, начиная тем самым новый круг движения вещества. Поэтому такие организмы-деструкторы (разрушители) называются еще редуцентами (от латинского слова редуцере - возвращать), или сапрофагами (от греческого слова сапрос - гнилой).
Пастбищная и детритная цепи в разных экосистемах присутствуют по-разному. Например, в лесу лишь небольшая часть зелени поступает в пищу консументам. Большая часть отмерших растений и их фрагментов поступает непосредственно к редуцентам. То есть лес считается экосистемой с преобладанием детритных цепей. В экосистеме гниющего пня пастбищная цепь вообще отсутствует. В то же время, например, в экосистемах поверхности моря практически все продуценты, представленные фитопланктоном, потребляются животными, а их трупы опускаются на дно, то есть уходят из данной экосистемы. В таких экосистемах, как говорят, преобладают пастбищные пищевые цепи или цепи выедания.
В любой экосистеме разные пищевые цепи не изолированы друг от друга, а переплетаются друг с другом в сложные пищевые (трофические) сети. Эти сети могут быть достаточно сложными и динамичными. Бывает очень трудно отнести какое-то животное к тому или иному уровню трофической сети. Ярким примером является человек, который питается как растениями, так и мясом животных. Тем не менее несмотря на некоторую условность деления трофической сети на уровни, в ней всегда присутствуют по крайней мере три уровня, обеспечивающие круговорот вещества в экосистеме: продуценты - консументы - редуценты.
Следует отметить, что с одного трофического уровня на другой передается не вся энергия данного уровня, а только та, которая накапливается в структуре организмов данного уровня. Основная часть энергии, усвоенной консументами с пищей, тратится на их жизнеобеспечение (дыхание). В сумме с неусвоенной пищей (экскременты) это составляет в среднем порядка 90 % от потребленной энергии. То есть энергия, накопленная в структурах организмов, а значит, передаваемая на следующий трофический уровень, в среднем составляет около 10 % от энергии, потребленной с пищей. Эта закономерность называется “правилом десяти процентов”.
На биосферу из космоса падает солнечный свет с энергией 2 кал/(мин.см2). Проходя через атмосферу, он ослабляется и в ясный летний день до поверхности Земли доходит не более 67 % его энергии, то есть 1,34 кал/(мин.см2), в пасмурный день ослабление еще существенней. За день к автотрофному слою поступает в среднем 300-400 кал/см2. Фотоактивная радиация, используемая при фотосинтезе, составляет порядка 40 % от поступившей солнечной радиации. Из нее растения связывают только около 1 % энергии. Только эта энергия, накопленная в органике растений, составляет первичную продукцию, которая затем может передаваться далее по пищевым цепям.
Из ограниченности количества поступающей энергии и правила десяти процентов следует, что все трофические цепи могут иметь только ограниченное количество уровней, как правило не больше 4-5. Количество живого вещества на каждом следующем уровне примерно на порядок меньше, чем на предыдущем.
Существует и еще одно следствие, очень важное для нашей цивилизации: с энергетической точки зрения потребление животной продукции, особенно с дальних уровней цепей питания, нецелесообразно. Примером могут служить пруды для спортивной ловли рыбы. Рыболову интересно вылавливать достаточно крупную рыбу, например окуня, который питается более мелкой рыбой. Поэтому для разведения окуней требуется водоем с большим количеством мелкой рыбы, питающейся зоопланктоном и мотылем, которые в свою очередь питаются фитопланктоном и его детритом. Пруд только с мелкой рыбой давал бы больше рыбы по биомассе, чем пруд с окунями, но человеку мелкие рыбы просто не интересны.
Особенно велики потери энергии при переходе от растений к травоядным животным. Поэтому с точки зрения роста народонаселения планеты энергетически наиболее выгодным является вегетарианство. Так, например, если мальчик весом 48 кг питался бы только мясом, то за год он съедал бы где-то 4,5 теленка, для выращивания которых требуется урожай люцерны с площади 4 га весом 8211 кг. Таков энергетический эквивалент питания ребенка.
При нормальном питании взрослый человек потребляет 80-100 кг мяса в год. При таком рационе уже невозможно обеспечить равноправие для 6 миллиардов людей планеты. При минимальном расходе мяса можно прокормить около 8 миллиардов людей. Переход всех людей на вегетарианство может обеспечить пищей приблизительно 15 миллиардов людей.
Эти цифры не зависят от успехов сельского хозяйства, а опираются только на данные по энергетике экосистем. Принципиальное ограничение наложено самим Солнцем. Правда, мы можем привлечь в сельское хозяйство дополнительные энергетические субсидии, в первую очередь от сжигания топлива и ядерных реакций. Собственно, это мы и делаем. Агросистемы - это яркий пример дополнительно субсидируемых экосистем. Здесь дополнительная энергия поступает в виде мышечных усилий человека и животных, работы машин, использующих горючее, орошения, внесения удобрений, пестицидов и т.п. Еще в прошлом веке Мальтус предупреждал, что уже 2 миллиарда людей Земля прокормить не в состоянии. Мы смогли превысить эту цифру только за счет энергетических субсидий в сельское хозяйство. Но этим мы неумолимо приближаем к себе другой аспект экологической катастрофы, которая пока еще нас мало тревожит, но первые ее предвестники уже говорят о себе в возросшей штормовой активности океанов, ослаблении стабильности антарктических ледниковых шельфов и т.п. Я имею в виду тепловую катастрофу. Сможем ли мы преодолеть и эту опасность? Очень сомнительно. Здесь запрет наложен одним из фундаментальнейших законов природы: принципом роста энтропии.
Для чего природе нужна столь сложная система передачи энергии? В чем положительная роль идеи сплошного поедания друг друга? Зачем нужны природе разрушители (деструкторы)? Во-первых, все консументы призваны вернуть вещество в круговорот. Без этого жизнь не смогла бы постоянно усложнять свои формы, то есть рано или поздно исчерпался бы лимит возможности роста энтропии. В рамках всей Вселенной это противоречит самим принципам ее существования. Таким образом, несмотря на то, что Земля - это “планета растений” (именно растения являются настоящими созидателями на планете), животные также необходимы для жизни биосферы. Во-вторых, чем сложнее трофическая сеть данной экосистемы, тем интенсивней круговорот веществ. Это облегчает поток энергии через экосистему. В-третьих, консументы - это не просто “пассивные едоки”. Удовлетворяя свои потребности в энергии, они регулируют всю экосистему, то есть являются основными звеньями механизмов гомеостаза экосистем. Причем реализуемые ими обратные связи могут быть не только отрицательными (выедание, то есть уменьшение биомассы предыдущего уровня трофической цепи), но и положительными. Так многие животные разными способами “ухаживают” за своими кормовыми растениями или как-то иначе способствуют их росту. Например, злаки, листья которых объедают кузнечики быстрее восстанавливаются, чем злаки с обрезанными листьями.
Дата: 2018-11-18, просмотров: 832.