Высшие растения поглощают соединения азота из почвы. Основным источником азотного питания для растений являются нитраты и аммиак. Изучение отдельных этапов превращения азотистых соединений, а также исследования, показавшие широкое распространение процессов реутилизации соединений азота, привели к представлению о круговороте азотистых веществ в растительном организме. Корневые системы растений хорошо усваивают нитраты, которые, поступая и корни растения, подвергаются ферментативному восстановлению до нитритов и далее до аммиака. Этот процесс происходит главным образом в корнях, однако этой способностью обладают и клетки листьев. Восстановление нитратов до аммиака идет через ряд этапов. На первом этапе нитраты восстанавливаются до нитритов при участии фермента нитратредуктазы:
NО3- + 2е -> NО2
Нитратредуктаза — это фермент с молекулярной массой 200—270 КДа, содержащий в своем составе ФАД, гем и молибден. Фермент локализован в цитозоле, где и протекает процесс восстановления нитратов до нитритов. Донором электронов при этой реакции у грибов является НАДФН, а у растений НАДН. В свою очередь поставщиком этих соединений являются процесс дыхания и отчасти световые реакции фотосинтеза. Именно поэтому восстановление нитратов тесно связано с дыхательным газообменом. Вместе с тем для нормального протекания процесса дыхания растение должно быть достаточно обеспечено углеводами. При искусственном снижении содержания углеводов (выдерживание растений в темноте) нитраты не восстанавливаются, а накапливаются во всех органах растения. При усиленном поступлении нитратов содержание углеводов падает. Интересно, что растения-нитратонакопители, например, некоторые среднеазиатские солянки, содержат мало углеводов и большое количество органических кислот. Показано, что нитратредуктаза относится к индуцибельным ферментам. Ее новообразование вызывается присутствием в среде нитратов. Вместе с тем накопление продукта реакции — нитритов репрессирует образование нитратредуктазы. Интересно, что фитогормоны цитокинины также индицируют синтез нитратредуктазы. На восстановление нитратов большое влияние оказывает свет. Прежде всего, на свету в процессе фотосинтеза образуются углеводы, необходимые для восстановления, а также для дальнейшего превращения нитратов. Вместе с тем для восстановления нитратов могут быть непосредственно использованы продукты, образующиеся в процессе нециклического фотофосфорилирования (НАДФН, АТФ). Свет влияет и на уровень фермента нитратредуктазы. Показано, что при низкой освещенности, дефиците Fe и Мо активность фермента снижается, и нитраты накапливаются в клетке. Восстановление нитратов стимулируется при освещении синим светом. Возможно, это связано с тем, что флавин, который входит в состав нитратредуктазы, поглощает синий свет и активируется им.
Второй этап — восстановление нитратов до аммиака катализируется ферментом нитритредуктазой:
NО2- + 6е -> NH4+
Нитритредуктаза — это фермент с молекулярной массой 60—70 КДа содержит в качестве простетической группы гем. Активность этого фермента значительно выше, чем нитратредуктазы. Нитритредуктаза локализована в хлоропластах листьев или пропластидах корней. Донором электронов в листьях служит восстановленный ферредоксин, который образуется при функционировании на свету ФС I. Нитриты образуются не только на промежуточной стадии восстановления нитратов. Они, как и нитраты могут поступать в растение из почвы. При этом нитриты также подвергаются восстановлению до аммиака при участии нитритредуктазы. Однако нитриты при накоплении в цитоплазме могут оказаться ядовитыми, поскольку фермент локализован в хлоропластах. Передвижение нитритов в хлоропласты стимулируется Са. При недостатке Са нитриты не восстанавливаются до аммиака и накапливаются в клетках. Установлено, что в высших растениях, так же как у прокариот и грибов, наряду с восстановлением нитратов до аммиака осуществляется и обратный процесс — окисление аммонийной формы азота в нитратную, что опровергает широко распространенное мнение об исключительно экзогенном происхождении нитратов в растениях (Б.А. Ягодин).
Процесс восстановления нитратов может осуществляться в листьях и корнях растений. Так, древесные растения, черника, люпин и другие восстанавливают нитраты преимущественно в корнях, а в листья транспортируют в органической форме. К видам, осуществляющим этот процесс в листьях, относятся свекла, дурнишник, хлопчатник. Однако большинство растений (злаки, бобовые, томаты, огурцы и др.) могут восстанавливать нитраты как в листьях, так и в корнях, что зависит от уровня снабжения нитратами. Важнейшим источником азотного питания является аммонийный азот. При этом он поступает в растения даже быстрее, чем нитраты. Более быстрое поглощение аммиака объясняется тем, что для его использования на построение органических веществ не требуется предварительного восстановления, которое необходимо при питании растений нитратами. Аммиак представляет собой основное и, по-видимому, единственное соединение, вовлекаемое в процессы азотного обмена. При этом аммиак может быть разного происхождения: непосредственно поступивший из почвы, образовавшийся в результате восстановления нитратов или в результате вторичного распада белка в стареющих органах и клетках. Важно отметить, что накопление аммиака в клетках, как растений, так и животных приводит к нежелательным последствиям и даже отравлению организма. Однако растения обладают способностью обезвреживать аммиак путем присоединения его к органическим кислотам с образованием амидов (глутамина, аспарагина). Этот процесс аналогичен обезвреживанию аммиака животными организмами в виде мочевины. Существует целая группа растений, накапливающая большое количество органических кислот и с их помощью обезвреживающая аммиак, образуя соли. Это позволило разделить растения на амидные, образующие амиды — аспарагин и глутамин, и аммиачные, образующие соли аммония. Изменяя рН клеточного сока, можно менять направление азотного обмена, превращать растения с амидным типом обмена в аммиачные и наоборот. Каковы же пути образования амидов в растениях? В процессе дыхания в качестве промежуточных продуктов образуются органические кислоты, в том числе а-кетоглутаровая и щавелевоуксусная. Эти кислоты в результате реакции прямого восстановительного аминирования присоединяют аммиак:
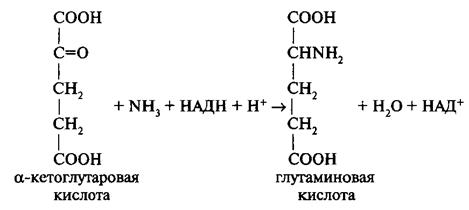
Реакция идет в две стадии. На промежуточном этапе образуется иминокислота. Катализируется реакция ферментом глутаматдегидрогеназой с активной группой НАД. Этот фермент локализован главным образом в митохондриях, так как именно в этих органеллах образуются органические кислоты и восстановленные никотинамидные коферменты, но может содержаться в цитозоле и хлоропластах. Аспарагиновая кислота образуется по аналогии с глутаминовой кислотой путем восстановительного аминирования щавелевоуксусной кислоты при участии фермента аспартатдегидрогеназы. На активность глутаминсинтетазы влияет присутствие катионов: Mg2+, Мп2+, Со2+, Са2+. Фермент обнаружен в цитозоле, но может находится и в хлоропластах. Образование аспарагина происходит аналогичным путем. Для образования амидов особенное значение имеет возраст растений. Как правило, чем моложе растение, тем больше его способность к образованию амидов. В более молодых органах (листьях) и даже в более молодых клетках одного и того же органа образование амидов идет интенсивнее. В пасоке и в гутте (сок гуттации) обычно присутствуют амиды. Это показывает, что аммиак, поступивший в растение, может преобразовываться в форму амидов в живых клетках корня. В тех случаях, когда не хватает углеводов или ослаблена интенсивность дыхания, амиды не образуются и накапливается аммиак. В результате может наступить отравление растений. Относительное количество образовавшегося аспарагина и глутамина и их роль различны в зависимости от вида растений и условий среды. Все же, по-видимому, образование аспарагина преобладает в том случае, когда происходит распад белков в семенах. В клетках корня и листа растущего растения идет, главным образом, образование глутамина. Таким образом, аспарагин — форма обезвреживания аммиака, образовавшегося на пути распада белка (регрессивная ветвь азотного обмена), тогда как глутамин — форма обезвреживания аммиака, используемого на пути синтеза белка (прогрессивная ветвь азотного обмена). Роль амидов в растении разнообразна. Это не только форма обезвреживания аммиака, это и транспортная форма азотистых соединений, обеспечивающая отток их из одного органа в другие. Наконец, чрезвычайно важно, что амиды и их непосредственные предшественники — глутаминовая и аспарагиновая кислоты — являются материалом для построения многих других аминокислот в процессах переаминирования, а также перестройки их углеродного скелета. Из 20 аминокислот, входящих в состав белка (протеиногенных), только три, как мы видим, могут образоваться в процессе прямого аминирования. Остальные аминокислоты образуются в процессе переаминирования и взаимопревращения. Каждая из аминокислот, образовавшихся путем прямого аминирования (глутаминовая, аспарагиновая и аланин), является предшественником целой группы аминокислот. Реакции переаминирования были открыты в 1937 г. отечественными биохимиками А. Е. Браунштейном и М.Г. Крицман. При этих реакциях аминогруппа от указанных аминокислот обменивается с кетогруппой любой кетокислоты с образованием соответствующей аминокислоты. Эти реакции катализируются специальными ферментами — аминотрансферазами и идут при участии кофермента пиридоксальфосфата (производное витамина В6):
R1CHNH2COOH + Р2СОСООН -> R1COCOOH + R2CHNH2OOH
Полученные путем переаминирования различные аминокислоты за счет перестройки углеродного скелета дают остальные аминокислоты. Таким образом, глутаминовая, аспарагиновая кислоты и их амиды являются как бы донорами аминогруппы. Д.Н. Прянишников называл их воротами, через которые должен пройти аммиак, для того чтобы включиться в остальные аминокислоты и белки. Растительный организм, в отличие от животного, обладает способностью синтезировать все необходимые ему аминокислоты из неорганических соединений. В этом также выражается его автотрофность. Аминокислоты могут образовываться в разных органах растений — в листьях, корнях, верхушках стебля. Некоторые аминокислоты образуются непосредственно в хлоропластах и здесь используются на образование белка. Наиболее интенсивно синтез белка происходит в меристематических и молодых развивающихся тканях. Интересно, что в отрезанных листьях синтез белка полностью прекращается. Это служит еще одним доказательством, что для синтеза белка нужен какой-то фактор, образующийся в корнях растений. Можно предположить, что это фитогормон, относящийся к группе цитокининов. Для нормального протекания синтеза белка в растительном организме нужны следующие условия: 1) обеспеченность азотом; 2) обеспеченность углеводами (углеводы необходимы как материал для построения углеродного скелета аминокислот и как субстрат для дыхания); 3) высокая интенсивность и сопряженность процесса дыхания и фосфорилирования. На всех этапах преобразования азотистых веществ (восстановление нитратов, образование амидов, активация аминокислот при синтезе белка и др.) необходима энергия, заключенная в макроэргических фосфорных связях (АТФ); 4) присутствие нуклеиновых кислот. ДНК необходима как вещество, в котором зашифрована информация о последовательности аминокислот в синтезируемой молекуле белка; мРНК — как агент, обеспечивающий перенос информации от ДНК в цитоплазму; тРНК — как обеспечивающая перенос аминокислот к рибосомам; 5) рибосомы — структурные единицы, где происходит синтез белка; 6) белки-ферменты — катализаторы синтеза белка (аминоацил-т-РНК-синтетазы); 7) ряд минеральных элементов (ионы Mg2+, Са2+и др.). Образованием белка заканчивается прогрессивная ветвь азотного обмена в растениях, которая преобладает главным образом в молодых растущих органах (первичный синтез белковых веществ). Однако в растениях идет и непрерывный распад белка. Опыты с использованием меченого азота 15N позволили исследователю Ф.В. Турчину подтвердить последовательность включения азота в различные соединения, постулированную схемой Прянишникова, и одновременно показать, что обновление белка происходит чрезвычайно быстро. За 48 ч до 60% белка организма синтезируется вновь. Белки распадаются до аминокислот и далее до аммиака. Аммиак вновь обезвреживается в виде амидов (аспарагин и глутамин). На основе этих соединений образуются аминокислоты. Это позволяет организму синтезировать новый набор аминокислот, который обеспечит построение иных белков со своим специфическим набором и последовательностью аминокислот (вторичный синтез белковых веществ). Анализируя факты, Д.Н. Прянишников подчеркнул, что аммиак — это альфа и омега азотного обмена в растениях, т. е. его начальный и конечный этапы. В условиях, обеспечивающих достаточно высокий уровень синтетических процессов, аммиак представляет собой прекрасный источник азотного питания для растений. Признание аммиака как источника азотного питания имеет не только теоретическое, но и практическое значение. Получение удобрений, содержащих аммиачные соли, — процесс более простой и дешевый по сравнению с удобрениями, где азот содержится в форме нитратов.
В целом, оценивая сравнительное физиологическое значение аммиачных и нитратных форм азотных удобрений, необходимо учитывать следующее:
1. Аммиак как источник азотного питания имеет то преимущество, что он быстрее поступает и быстрее используется растением. Однако во избежание аммиачного отравления растения необходимо присутствие в нем достаточного количества углеводов. В силу этого применение аммиачных удобрений в рядки для культур с мелкими семенами не рекомендуется.
2. При внесении различных форм азота важно учитывать реакцию питательной среды: в слабокислой почве лучше поглощаются нитраты, в нейтральной преимущество имеет аммонийная форма. Поэтому для усвоения аммонийного азота большое значение имеет одновременное достаточное снабжение кальцием.
3. Накопление нитратов для растительного организма не представляет опасности. Однако при высоких дозах и накоплении нитратов в клетках растений они становятся весьма опасными для животных организмов, в том числе и для человека. Попадая в желудок, они могут восстанавливаться до нитритов и далее при взаимодействии с вторичными аминами образовывать нитрозамин — вещество, обладающее сильным мутагенным действием. При этом необходимо учитывать, что повышенной способностью к накоплению нитратов при внесении удобрений обладают кормовые злаковые травы и некоторые овощные культуры. Считается, что предельно допустимая норма содержания нитратов в овощах не должна превышать 290—300 мг на 1 кг сырой массы. Предельно допустимое количество (ПДК) нитратов для человека составляет 500 мг в день. Это еще раз показывает необходимость строгого нормирования внесения питательных веществ с удобрениями.
4. Наконец, аммиачные и нитратные удобрения оказывают различное влияние на процессы обмена веществ и, как следствие, на качество урожая. При внесении аммиачных форм в растении накапливаются восстановленные соединения, тогда как при нитратных — окисленные. Так, было показано, что внесение аммиачных форм удобрений под махорку позволяет усилить накопление в ней никотина, тогда как внесение нитратов вызывает преимущественное образование органических кислот (А.В. Владимиров).
Опыты, проведенные в стерильных условиях, показали, что в качестве источника азотного питания могут быть использованы растениями и растворимые органические соединения (аминокислоты, амиды и мочевина). Однако их содержание в почве, как правило, очень мало. Для некоторых растений с уклоняющимся типом питания (паразиты, полупаразиты, сапрофиты, несекомоядные растения) источником питания может служить органический азот.
Дата: 2018-12-21, просмотров: 1093.