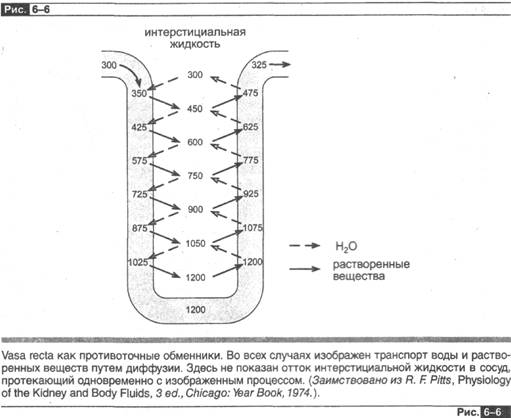
Кровообращение мозгового слоя обладает уникальной особенностью, без которой не смогла бы функционировать прртивоточная множительная система, она обусловлена шпилькообразной формой прямых сосудов, особым анатомическим строением сосудов мозгового слоя — vasa recta , которые проходят параллельно петлям Генле и собирательным трубкам мозгового вещества. Зададим себе вопрос: что произойдет с градиентом в интерстициальном 'пространстве мозгового слоя, если бы последний снабжался кровью только обычных капилляров? По мере того как плазма с обычной осмолярностью, равной 300 мосм/л, поступает в очень концентрированную окружающую среду мозгового слоя, происходит диффузия хлорида натрия в капилляры и отток воды из них. Вследствие этого величина интерстициального градиента должна стремиться к нулю. Однако наличие шпилькообразной структуры обусловливает ряд последовательных событий, которые изображены на рис. 6-6. Кровь поступает в сосудистую петлю, имея осмо-лярность 300 мосм/л; она течет вниз по петле капилляров в глубь мозгового слоя, хлорид натрия диффундирует внутрь сосуда, а вода — из него. Преодолев вершину шпильки — перегиб петли, кровь течет далее вверх по восходящей части сосудистой петли, где идет процесс, обратный описанному.
Таким образом, сосудистая петля функционирует как Противоточный обмен- ник, который препятствует исчезновению градиента. Обратите внимание, сосуд рег зе функционирует пассивно; т. е. не он создает градиент в мозговом слое, он его только сохраняет. Благодаря пассивной роли он получил название обменника. Сравните его функцию с деятельностью петли Генле, которая создает градиент, и станет ясно, почему она названа умножителем.
В конечном счете следует отметить, что шпилькообразная структура кровеносных сосудов сводит к минимуму потери растворенных веществ, а также воды из интерстициального пространства посредством диффузии. Тем не менее это не предотвращает переход части интерстициальной жидкости мозгового слоя в капилляры, что происходит вторично в соответствии с действием сил Старлинга. Результатом процесса перехода хлорида натрия и воды, поступающих в интерсти-циальное пространство из петель Генле и собирательных трубок, является удаление указанных веществ из этих частей канальца и поддержание стационарного градиента в этой зоне почки.
Однако vasa recta не являются совершенными противоточными обменниками, они удаляют накопившиеся в результате диффузии растворенные вещества э несколько большем количестве, чем воду. Это объясняет, почему мозговой кровоток намного менее интенсивен, чем корковый. Однако он должен быть достаточно выражен, чтобы уносить с током крови реабсорбированную воду и хлорид натрия, как это было описано в предыдущем параграфе. Соответственно изменения кровотока в мозговом слое — и слишком большие, и слишком маленькие — будут уменьшать интерстициальный градиент и, следовательно, степень гиперосмоляр-
ности мочи14
134
Основные почечные процессы , касающиеся транспорта натрия , хлора и воды
Резюме
На рис. 6-7 изображены в обобщенном виде описанные изменения объема и осмолярности канальцевой жидкости по мере ее продвижения по нефрону.
1. Примерно 65 % натрия, хлора и воды реабсорбируется в проксимальном ка-
, нальце, но жидкость остается йзоосмотичной.
2. В петле вода реабсорбируется из нисходящей части, но гораздо большие ко
личества хлорида натрия реабсорбируются из восходящей части, поэтому в
дистальный извитой каналец поступает гипоосмотичная жидкость.
3. В дистальном извитом канальце жидкость по-прежнему остается гипоосмо-
тичной, при этом вода реабсорбируется в минимальном количестве или во
обще не реабсорбируется. Из сказанного следует, что восходящие части пе
тель Генле и дистальный извитой каналец функционируют как сегменты, где
происходит разведение жидкости.
4. Присутствие или отсутствие АДГ имеет значение только для функции соби
рательных трубок. При почти полном отсутствии АДГ в плазме очень не
большое количество воды реабсорбируется из собирательных трубок, поэто
му указанные сегменты способствуют разведению мочи, соответственно об
разуется очень большой объем разведенной мочи.
135
| максимальное содержание АДГ |
|
|
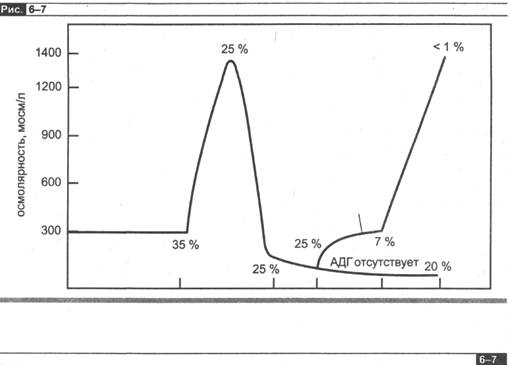
Осмолярность канальцевой жидкости и процент профильтровавшейся воды, остающейся в различных отделах канальца. (Цифры, касающиеся процента реабсорбции воды в каждом сегменте канальца, заимствованы из табл. 6-3.)
Рис .
5. При высокой концентрации АДГ в плазме в собирательных трубках реаб-
сорбция воды велика. В конечной части собирательных трубок коркового ве
щества жидкость вновь становится изоосмотичной плазме корковой зоны.
Большее количество оставшейся воды реабсорбируется в собирательных
трубках мозгового слоя, поэтому образуется небольшой объем высококон
центрированной мочи.
Следует обратить пристальное внимание на некоторые моменты, имеющие особое значение:
6. Значительные количества воды могут быть экскретированы даже при усло
вии, что моча практически будет свободна от натрия и других растворенных
веществ; это результат водного диуреза, который наблюдается при низкой
концентрации АДГ в плазме.
6. Экскреция значительных количеств натрия или других растворенных ве
ществ всегда приводит к экскреции значительного объема воды, этот процесс
называют салурезом. Он основан на том, что вода может быть реабсорбиро-
вана только в том случае, если вначале будет реабсорбировано растворенное
в ней вещество. Почти все лекарственные средства (диуретики), применяе
мые в клинической практике для увеличения мочеотделения, действуют
благодаря угнетению (прямо или косвенно) одного или двух процессов
транспорта, осуществляющих реабсорбцию натрия и/или хлора (см. Прило
жение В, где резюмированы данные, касающиеся действия этих лекарств).
Немедикаментозный вариант диуреза представляет собой осмотический ди
урез, который описан в разделе,'касающемся проксимального канальца.
136
Основные почечные процессы,
натрия, хлора и воды
Рассмотрев в этой главе основные процессы транспорта в почке натрия, хлора и воды, мы обратимся к рассмотрению механизмов, которые их регулируют с тем, чтобы обеспечить водно-солевой гомеостаз.
Вопросы для изучения: 33—40.
Примечания
1 Вероятно в исключительных обстоятельствах хлор макет секретироваться. Это наблюдается в дистальном извитом канальце и собирательных 1ру(яиа. коркового вещества (см. статьи Сге^ег, Коерреп и 51ап1;оп).
г \
2В действительности помимо бикарбоната данный переносчик способен активно транспортировать и небольшое количество натрия через баэовяерадьную мембрану. При сопряжении 3 ионов НСО3 с 1 атомом Ка, в чем нас убежяиот представленные факты, движение по градиенту столь значительного количества бикарбоната может инициировать движение натрия против градиента. Но количество перенесенного натрия очень мало по сравнению с тем, который транспортируется Ыа,К-АТФаэон.
3 Эти три вида эпителия являются типичными представителями, распространенными в природе. Говоря о разнице проницаемости, обычно имеют в виду разницу проницаемости люминальных мембран, которые служат барьером, регулирующим скорость переноса (поскольку проницаемость базолатеральной мембраны обычно выше, чем апикальной) (см. ЗсЬа&г и соавт.).
4В дополнение к котранспортерам и антипорту, которые действуют в проксимальном канальце (см. табл. 6-1), в данном сегменте также найдены натриевые каналы. Доля участия этих каналов в настоящее время неизвестна (см. РаЬпег).
5Выраженная реабсорбция хлора, зависящая от трансканальцевой разницы его концентрации, в значительной степени определяет появление небольшого положительного со стороны просвета трансканальцевого электрического потенциала, в данном отделе нефрона. Этот потенциал вызывает небольшую парацеллюлярную реабсорбцию натрия. В то же время, как отмечено в главе 4, в начальном участке проксимального канальца имеет место небольшой отрицательный со стороны просвета трансканальцевый потенциал, что облегчает парацеллюлярную секрецию натрия. В итоге можно считать потенциал проксимального канальца нулевым и сосредоточить внимание на активном транспорте натрия (и на трансканальцевом концентрационном градиенте, обеспечивающем пассивный перенос хлора). Другое соображение, по которому мы игнорировали пассивную реабсорбцию натрия в конечном отделе проксимального канальца, заключается в том, что если вы вернетесь к началу изложения, то увидите, что этот тип реабсорбции имеет малое значение для активной реабсорбции натрия против градиента в начальном отделе проксимального канальца: реабсорбция в начальном отделе проксимального канальца -> реабсорбция воды в начальном отделе проксимального канальца -> концентрирование в просвете хлора -> диффузия хлора через стенку в конечных отделах проксимального канальца -» положительная разность потенциалов со стороны просвета -> пассивная реабсорбция натрия. В конечном счете мы не рассматривали также реабсорбцию как натрия, так и хлора, осуществляемую механизмом переноса веществ вместе с растворителем, поскольку, как описано в следующем разделе, реабсорбция воды тоже зависит от активной реабсорбции натрия. (Для более подробного анализа механизма переноса веществ вместе с растворителем и реабсорбции натрия в проксимальном канальце см. статьи ЗсЬа&г с соавт. и ОДш&еш.)
г *
6Слово «существенно» означает, что величины осмолярности клубочков ого фильтрата и интерстициальной жидкости не равны. В интеретициальной жидкости почки находится
137
белок в значительной концентрации, и разница в 1—2 мосм/л, обусловленная присутствием белка, влечет за собой реабсорбцию некоторого количества воды (см. ЗсЬа&г с соавт.).
7В действительности этого может не произойти в существенных размерах, либо действие будет полностью нейтрализовано, поскольку интерстициальное пространство велико, и очень значительный объем крови, протекающей по перитубулярным капиллярам, оказывает стабилизирующее действие на его состав. Другой потенциальный фактор, влияющий на значительную разницу осмолярности жидкости в просвете канальца и осмолярности жидкости в интерстициальном пространстве, может зависеть от величины коэффициента отражения разных веществ, растворенных в канальцевой жидкости; обсуждение этого вопроса я не буду здесь приводить.
о
8У некоторых видов экспериментальных животных Ка,С1-котранспортер имеется не в клетках дистального извитого канальца, а в клетках следующего сегмента — связующего отдела. Данных, касающихся человека, пока нет.
9Рецепторы, функционирующие в собирательной трубке, относятся к типу У2. Рецепторы V, локализованы на гладкой мускулатуре сосудов и они опосредуют сосудосуживающий эффект АДГ.
10Такие каналы существуют и в более проксимальных отделах канальцев, но не регулируются АДГ (см. Уегктап).
11 АДГ осуществляет местное влияние с помощью механизма обратной отрицательной связи на собственный эффект. АДГ усиливает в мозговом веществе почки синтез и освобождение простагландинов, которые затем тормозят действие АДГ, вмешиваясь в стимулированный АДГ-синтез циклического АМФ. Соответственно нарушенный сцнтез простагландинов (усиленный или сниженный) может влиять на изменение канальцевой реакции на АДГ, что отмечается при некоторых заболеваниях почек или при лечении лекарствами, которые блокируют синтез простагландинов. На реакцию клеток на АДГ влияют не только простагландины. Например, кортикостероиды вмешиваются в ряд этапов эффекта АДГ; поэтому у больных с недостаточностью надпочечников имеется тенденция к гиперреактивности к АДГ.
12Нисходящая часть петли Генле не является полностью непроницаемой для натрия и хлора. Поэтому некоторая часть этих ионов диффундирует в петлю одновременно с водой из петли. Для простоты изложения мы не будем вдаваться в детали этого процесса (см. ^ат^5оп, Сепп§ и Коу с соавъ).
Л О
13 Как описывалось ранее, натрий и хлор также активно реабсорбируются из собирательных трубок. Этот процесс способствует уменьшению потери хлорида натрия с мочой, но он не является очень значимым компонентом противоточной системы; т. е. его вклад в количество растворенных веществ, поступающих в интерстициальное пространство, невелик по сравнению с количеством веществ, реабсорбируемых в восходящей части петли Генле.
14 Как описано в главе 2, кровоток в мозговом веществе может в значительной степени подвергаться независимой регуляции. Вполне возможно, что главной функцией такой регуляции является действие на процесс концентрирования мочи при участии противоточной системы (см. статьи Спои с соавт. и РаПопе с соавт. в списке литературы к главе 2).
Глава 7
Регуляция экскреции натрия и воды :
Дата: 2018-09-13, просмотров: 851.