ОБЩИЕ ПРЕДСТАВЛЕНИЯ ОБ ИМПРИНТИНГЕ
Оба родителя передают потомкам совершенно идентичные гены, но в гаметогенезе они могут быть по-разному маркированы, вследствие чего их действие будет неодинаковым. Геномный импринтинг (ГИ) относят к эпигенетическим явлениям, подчеркивая этим наследуемость изменений генной активности, обусловленной родительским происхождением, а не структурной перестройкой генетического материала. Таким образом, ГИ можно определить как эпигенетический механизм регуляции экспрессии гомологичных генов в процессе развития организма в зависимости от родительского происхождения гена, хромосомы или генома. Импринтированные гены дифференциально экспрессируются в зависимости от материнского или отцовского происхождения, т.е. в диплоидной клетке млекопитающих они экспрессируются только с одного аллеля.
Нарушением, демонстрирующим импринтинг всего генома у человека, считают истинный пузырный занос - быстро малигнизирующуюся ткань хориона абортивного плодного пузыря. Цитогенетическими и молекулярно-генетическими методами было показано, что ткань пузырного заноса - продукт андрогенеза, т.е. возникает при оплодотворении яйцеклетки, лишенной хромосом матери, двумя сперматозоидами 22+X. Несмотря на существование полноценного диплоидного набора, ранний эмбриогенез таких андрогенетических зигот протекает аномально, ткани собственно эмбриона вообще не формируются, но бурно разрастается трофобласт. В случае двойного набора материнских хромосом (гиногенетическая зигота) развивается тератома - эмбриональная опухоль, включающая все три эмбриональных слоя и не содержащая плацентарную ткань. Следовательно, геном отца обеспечивает развитие плаценты, в то время как геном матери - раннее развитие эмбриональных структур. Будучи генетически импринтированным, только материнский или только отцовский геном не в состоянии обеспечить нормальное эмбриональное развитие.
Аномалии развития, свидетельствующие о ГИ, обнаружены в случаях триплоидии у человека. Триплоидный плод - андроид (2p+1m) - характеризуется большой головой, маленьким веретенообразным телом, синдактилией, отставанием в росте и развитии. Формируется большая кистозная плацента - частичный пузырный занос. У гиноида (2m+1p) плацента недоразвита, эмбрион не развивается и представлен недифференцированной клеточной массой. Это также подтверждает неэквивалентность функционирования мужского и женского генома.
На хромосомном уровне импринтинг был обнаружен при получении мышиных транслокационных гибридов, которые имели в хромосомном наборе фрагменты или целые хромосомы одного (материнского или отцовского) происхождения - так называемая однородительская дисомия (ОРД). Термины «однородительская дисомия» или «изодисомия » предложены Эриком Энжелом в 1980 г. и обозначают типы анеуплоидии в половых клетках млекопитающих, которые свидетельствуют о том, что у диплоидного потомка обнаруживают два локуса от одного родителя. ОРД по целым хромосомам или их фрагментам обнаружены при анализе наследственных нарушений у человека и будут описаны ниже. Сейчас в геноме человека известно около 150 импринтированных генов, имеющих тканеспецифическую моноаллельную экспрессию, хотя предполагают, что их может быть не менее 200.
МЕХАНИЗМЫ ГЕНОМНОГО ИМПРИНТИНГА
Согласно современным представлениям, в основе геномного импринтинга у млекопитающих лежат специфические структурно-молекулярные изменения некоторых хромосомных участков, возникающие во время формирования мужских и женских половых клеток. Они приводят к стойким различиям экспрессии отцовских и материнских генов у потомства. Предполагают, что импринтинг модифицирует структуру гена так, что регуляторные факторы, действие которых манифестирует в клетке позднее, распознают импринтированные (меченые, видоизмененные каким-либо образом) материнские или отцовские аллели и избирательно их инактивируют.
Совокупность модификаций, которые по-разному маркируют родительские аллели и обеспечивают моноаллельный характер экспрессии импринтированных генов на хромосомах отцовского или материнского происхождения, называют эпигенотипом (импринтом).
При изучении механизмов импринтинга центральным считают вопрос о молекулярном происхождении тех эпигенетических модификаций, в которых записана информация (память) о функциональной активности генов на материнских и отцовских хромосомах. Существующие представления об эпигенетических модификациях были получены в результате поиска молекулярных различий между материнскими и отцовскими копиями импринтированных генов. Наиболее изученная эпигенетическая модификация, которой отводят основную роль в обеспечении процессов импринтинга, - специфическое метилирование цитозиновых остатков ДНК. Важнейшие особенности метилирования ДНК: стабильное сохранение в ряду многих поколений клеток и регуляция экспрессии генов.
В настоящее время установлено, что все известные импринтированные гены содержат области различного (дифференциального) метилирования на двух родительских хромосомах, причем эти различия обязательны для их моноаллельной экспрессии. Необходимость процесса метилирования в обеспечении механизмов импринтинга была доказана в экспериментах на мышах с дефектным геном ДНК-метилтрансферазы, у которых был утрачен моноаллельный характер экспрессии импринтированных генов H19, Igf2, Igf2r и Snrpn. У мышей, нокаутированных по гену метилтрансферазы, эмбриогенез останавливается на уровне восьми сомитов.
Гиперметилирование гена обычно подавляет его транскрипционную активность и в большинстве случаев происходит на неактивных аллелях. Дифференциальное метилирование родительских аллелей, как правило, происходит внутри или рядом с CG-богатыми районами, которые нередко содержат разные типы прямых повторов. Установлены некоторые характерные особенности этих повторов: 1) между ними нет гомологии; 2) длина единицы повтора может быть различной; 3) возможно любое расположение повторяющихся последовательностей по отношению к гену (рядом с геном с 5’-стороны, в 3’-нетранслируемой области, внутри интрона или в кодирующей части).
Дифференциально метилированные районы в некоторых случаях перекрываются с районами, с которых транскрибируются некодирующие и антисмысловые РНК, причем транскрипты включают блоки тандемных повторов. Такие РНК осуществляют регуляторные функции в импринтированных районах.
Функциональная активность импринтированных генов, помимо метилирования, в значительной степени определяется структурной организацией хроматина, что можно подтвердить явлениями асинхронной репликации и разной чувствительностью родительских аллелей к нуклеазной обработке. Исследование структуры хроматина в критическом районе хромосомы 15 показало, что промотор и первый экзон SNRPN гиперчувствительны к нуклеазам на отцовской хромосоме. Это свидетельствует о транскрипционно активном состоянии гена и о том, что эта часть центра импринтинга (ЦИ) контролирует отцовский эпигенотип.
Одно из ведущих направлений в изучении механизмов импринтинга - наблюдение за изменением эпигенетических свойств отцовских и материнских аллелей на протяжении последовательных стадий онтогенетического развития и анализ влияния эпигенотипа на функциональную активность импринтированных локусов. В результате многочисленных экспериментов на мышах была подробно изучена динамика метилирования и деметилирования генома в ходе онтогенеза. В частности, было показано, что метилирование импринтированных, равно как и неимпринтированных, генов теряется («стирается») на самых ранних стадиях развития гамет, а именно в примордиальных герминальных клетках эмбриона (рис. 9). Во всех экспериментах с такими клетками зародышевого пути отмечена биаллельная экспрессия импринтированных локусов. Следовательно, на этом этапе эпигенетические различия между отцовскими и материнскими аллелями отсутствуют. Это подтверждают эксперименты, в которых после слияния зрелых лимфоцитов тимуса с клетками зародышевой линии на ранней стадии развития происходило массивное деметилирование всего генома лимфоцита, включая импринтированные и неимпринтированные локусы.
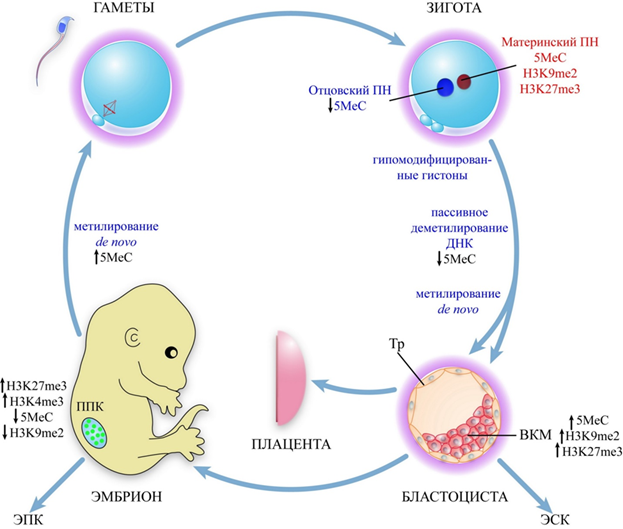
Рисунок 9. Динамика эпигенетических модификаций генома на ранних этапах эмбрионального развития: ПН — пронуклеус; ВКМ — внутренняя клеточная масса; ТЭ — трофэктодерма; ЭСК — эмбриональные стволовые клетки; ЭГК — эмбриональные герминальные клетки; ↑ — повышение; ↓ — снижение.
Установление метилирования в импринтированных локусах происходит на последующих этапах дифференцировки гамет. В овогенезе установление нового импринта происходит на стадии роста овоцита 1-го порядка, но полностью заканчивается перед овуляцией. Срок установления мужского эпигенотипа в сперматогенезе окончательно не выяснен, но считается, что это происходит в сперматоцитах 1-го порядка до наступления/во время профазы первого деления мейоза. Остается неясным механизм распознавания в отцовском и материнском гаметогенезе тех последовательностей, которые должны быть по-разному метилированы. В этом отношении представляет интерес обнаружение двух альтернативных способов сплайсинга 5’-экзонов гена DNMT1, один из которых реализуется в овогенезе, а второй - в сперматогенезе. Оказалось, что в сперматоцитах 1-го порядка на стадии профазы первого деления мейоза содержится та сплайс-форма DNMT1-мРНК, которая не транслируется и не дает белкового продукта. Овоцитспецифический сплайсинг гена DNMT1 сопровождается продукцией укороченных с N-конца молекул фермента, которые в большом количестве присутствуют в растущем овоците 1-го порядка и специфически метилируют будущие материнские аллели импринтированных генов. Возникает вопрос, каким образом в сперматогенезе происходит метилирование импринтированных локусов. Возможно, это происходит до наступления профазы первого деления мейоза с участием соматической изоформы DNMT1 или его осуществляют другие, пока неизвестные типы ДНК-метилтрансфераз или особая форма DNMT1.
После оплодотворения, между стадией 8-клеточного эмбриона и бластоцистой, у мыши происходит общее массивное деметилирование геномной ДНК, к которому устойчивы лишь некоторые импринтированные локусы. Метилирование ДНК начинается после имплантации эмбриона и продолжается в ходе гаструляции. Таким образом, аллельспецифический характер метилирования ДНК в ходе эмбриогенеза формируется постепенно, поэтому процессы импринтинга и метилирования могут носить многоступенчатый характер и приурочены к определенной стадии развития.
Хотя роль метилирования в обеспечении аллельспецифической экспрессии генов несомненна, остается неясным, служит оно первичным эпигенетическим сигналом, который стирается и устанавливается в гаметогенезе, или представляет некий вторичный процесс по отношению к более ранней стадии импринтинга и необходим лишь для поддержания ранее установленного импринта. Таким образом, механизмы геномного импринтинга, несмотря на значительное количество данных и моделей регуляции, все еще требует новых экспериментов, наблюдений и гипотез.
Дата: 2019-12-10, просмотров: 567.