В ЦНС наиболее быстро потенциал действия проводят миелинизированные нервные волокна. Но очень короткие участки этих волокон, называемые перехватами Ранвье, покрыты обычной клеточной мембраной. Таким образом, перехваты Ранвье - это области прерыва между соседними миелинизирующими клетками (обнаженная часть аксона между двумя соседними оболочками)
В межперехватных участках мембрана образует вокруг клетки многослойную оболочку, которая значительно увеличивает сопротивление мембраны. Возбуждение перескакивает от одного перехвата к следующему, без потери времени на межперехватных участках ( сальтаторное проведение ).
В периферической нервной системе функции олигодендроглии выполняют шванновские клетки нейроэктодермального происхождения. Они отличаются от олигодендроглии тем, что охватывают обычно только один участок отдельного аксона . Длина такого охвата не превышает 1 мм. Между отдельными шванновскими клетками формируются перехваты Ранвье
13 Распространяющийся потенциал действия, бездекрементное проведение, закон “всё или ничего”
Рис. 1 Фазы ПД
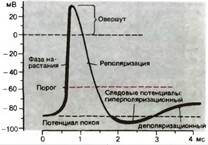
Потенциал действия - это электрический компонент нервного импульса, характеризующий изменения электрического заряда (потенциала) на локальном участке мембраны во время прохождения через него нервного импульса (от -70 до +30 мВ и обратно).
восходящая ветвь графика:
1. потенциал покоя – исходное обычное поляризованное электроотрицательное состояние мембраны (–70 мВ);
2. нарастающий локальный потенциал – пропорциональная раздражителю деполяризация;
3. критический уровень деполяризации (–50 мВ) – резкое ускорение деполяризации (за счет самораскрытия натриевых каналов), с этой точки начинается спайк – высокоамплитудная часть потенциала действия;
4. самоусиливающаяся круто нарастающая деполяризация;
5. переход нулевой отметки (0 мВ) – смена полярности мембраны;
6. «овершут» – положительная поляризация (инверсия, или реверсия, заряда мембраны);
7. пик (+30 мВ) – вершина процесса изменения полярности мембраны, вершина потенциала действия.
Нисходящая ветвь графика:
8. реполяризация – восстановление прежней электроотрицательности мембраны;
9. переход нулевой отметки (0 мВ) – обратная смена полярности мембраны на прежнюю, отрицательную;
10. переход критического уровня деполяризации (–50 мВ) – прекращение фазы относительной рефрактерности (невозбудимости) и возврат возбудимости;
11. следовые процессы (следовая деполяризация или следовая гиперполяризация);
12. восстановление потенциала покоя – норма (–70 мВ).
Итак, сначала – деполяризация, затем – реполяризация. Сначала – утрата электроотрицательности, затем – восстановление электроотрицательности.
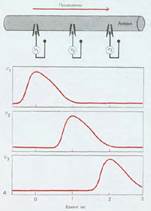 Рис.2 Бездекрементное проведение ПД
Рис.2 Бездекрементное проведение ПД
У высших животных благодаря прежде всего наличию миелиновой оболочки и совершенства метаболизма в нервном волокне возбуждение проходит, не затухая, бездекрементно. Этому способствуют наличие на всем протяжении мембраны волокна равного заряда и быстрое его восстановление после прохождения возбуждения.
В миелиновых волокнах возбуждение охватывает только участки узловых перехватов, т. е. минует зоны, покрытые миелином. Такое проведение возбуждения по волокну называется сальтаторным (скачкообразным). В узловых перехватах количество натриевых каналов достигает 12 000 на 1 мкм , что значительно больше, чем в любом другом участке волокна. В результате узловые перехваты являются наиболее возбудимыми и обеспечивают большую скорость проведения возбуждения. Время проведения возбуждения по миелиновому волокну обратно пропорционально длине между перехватами.
Проведение возбуждения по нервному волокну не нарушается в течение длительного (многочасового) времени. Это свидетельствует о малой утомляемости нервного волокна. Считают, что нервное волокно относительно неутомляемо вследствие того, что процессы ресинтеза энергии в нем идут с достаточно большой скоростью и успевают восстановить траты энергии, происходящие при прохождении возбуждения.
В момент возбуждения энергия нервного волокна тратится на работу натрий-калиевогонасоса. Особенно большие траты энергии происходят в перехватах Ранвье вследствие большой плотности здесь натрий-калиевых каналов.
Нервный ствол образован большим числом волокон, однако возбуждение, идущее по каждому из них, не передается на соседние. Эта особенность проведения возбуждения по нерву носит название закона изолированного проведения возбуждения по отдельному нервному волокну. Возможность такого проведения имеет большое физиологическое значение, так как обеспечивает, например, изолированность сокращения каждой нейромоторной единицы.
Способность нервного волокна к изолированному проведению возбуждения обусловлена наличием оболочек, а также тем, что сопротивление жидкости, заполняющей межволоконные пространства, значительно ниже, чем сопротивления мембраны волокна. Поэтому ток, выйдя из возбужденного волокна, шунтируется в жидкости и оказывается слабым для возбуждения соседних волокон. Необходимым условием проведения возбуждения в нерве является не просто его анатомическая непрерывность, но и физиологическая целостность. В любом металлическом проводнике электрический ток будет течь до тех пор, пока проводник сохраняет физическую непрерывность. Для нервного «проводника» этого условия недостаточно: нервное волокно должно сохранять также физиологическую целостность. Если нарушить свойства мембраны волокна (перевязка, блокада новокаином, аммиаком и др.), проведение возбуждения по волокну прекращается. Другим свойством, характерным для проведения возбуждения по нервному волокну, является способность к двустороннему проведению. Нанесение раздражения между двумя отводящими электродами на поверхности волокна вызовет электрические потенциалы под каждым из них.
Закон “ все или ничего”.
При допороговых раздражениях клетки в ткани ответной реакции не возникает. При пороговой силе раздражителя развивается максимальная ответная реакция, поэтому увеличение силы раздражения выше пороговой не сопровождается ее усилением. В соответствии с этим законом реагирует на раздражения одиночное нервное и мышечное волокно, сердечная мышца.
Со временем была установлена и относительность этого закона. Оказалось, что “все” зависит от функционального состояния ткани (охлаждение, исходное растяжение мышцы и т. д.).
С появлением микроэлектродной техники было установлено еще одно несоответствие: подпороговое раздражение вызывает местное, нераспространяющееся возбуждение, следовательно, нельзя говорить, что допороговое раздражение не дает ничего.
Процесс развития возбуждения подчиняется этому закону с уровня критической деполяризации, когда запускается лавинообразное поступление ионов K+ в клетку.
Дата: 2019-12-10, просмотров: 643.