Через фосфолипидный бислой могут диффундировать различные вещества, причем степень проницаемости (Р), т. е. способность клеточной мембраны пропускать эти вещества, зависит от разности концентраций диффундирующего вещества по обе стороны мембраны.
Именно ионные каналы обеспечивают два важных свойства мембраны: селективность и проводимость.
Селективность, или избирательность, канала обеспечивается его особой белковой структурой. Большинство каналов являются электроуправляемыми, т. е. их способность проводить ионы зависит от величины мембранного потенциала.
Рассмотрим принцип работы ионных каналов на примере натриевого канала. Полагают, что в состоянии покоя натриевый канал закрыт. При деполяризации клеточной мембраны до определенного уровня происходит открытие m-активационных ворот (активация) и усиление поступления ионов Na+ внутрь клетки. Через несколько миллисекунд после открытия m-ворот происходит закрытие п-ворот, расположенных у выхода натриевых каналов (инактивация)
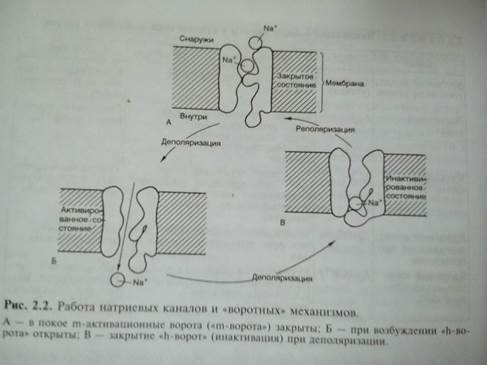
Свойство проводимости различных каналов неодинаково. В частности, для калиевых каналов процесс инактивации, как для натриевых каналов, не существует. Имеются особые калиевые каналы, активирующиеся при повышении внутриклеточной концентрации кальция и деполяризации клеточной мембраны. Активация калий-кальцийзависимых каналов ускоряет реполяризацию, тем самым восстанавливая исходное значение потенциала покоя.
Активация кальциевых каналов обеспечивается деполяризацией клеточной мембраны, например входящим натриевым током. Процесс инактивации кальциевых каналов достаточно сложен. С одной стороны, повышение внутриклеточной концентрации свободного кальция приводит к инактивации кальциевых каналов. С другой стороны, белки цитоплазмы клеток связывают кальций, что позволяет поддерживать длительное время стабильную величину кальциевого тока, хотя и на низком уровне; при этом натриевый ток полностью подавляется.
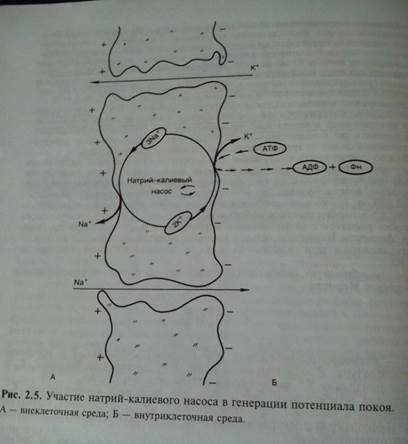
Известно, что в невозбужденном состоянии клеточная мембрана высокопроницаема для ионов калия и малопроницаема для ионов натрия.
Важнейшим механизмом, поддерживающим низкую внутриклеточную концентрацию ионов Na+ и высокую концентрацию ионов К+, является натрий-калиевый насос (рис. 2.9). Известно, что в клеточной мембране имеется система переносчиков, каждый из которых связывается с 3 находящимися внутри клетки ионами Na+ и выводит их наружу. С наружной стороны переносчик связывается с 2 находящимися вне клетки ионами К+, которые переносятся в цитоплазму. Энергообеспечение работы систем переносчиков обеспечивается АТФ.
Деполяризующий толчок тока приводит к активации натриевых каналов и увеличению натриевого тока.
Аналогичный эффект удавалось получить при добавлении в перфузионный раствор ТТХ (тетродотоксин) — специфического блокатора натриевых каналов
На пике потенциала действия проводимость мембраны для ионов натрия (gNa+) начинает быстро снижаться. Этот процесс называется инактивацией. Скорость и степень натриевой инактивации зависят от величины мембранного потенциала, т. е. они потенциалзависимы.
При постепенном уменьшении мембранного потенциала до —50 мВ (например, при дефиците кислорода, действии некоторых лекарственных веществ) система натриевых каналов полностью инактивируется и клетка становится невозбудимой.
Еще рекомендую почитать методичку «Физиология возбудимых тканей» про ионные каналы.
10. Роль ионов Ca2+ , поверхностных зарядов в регуляции ионной проницаемости мембран
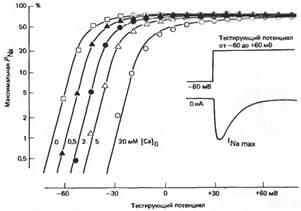
ЗАВИСИМОСТЬ Na+ -ПРОНИЦАЕМОСТИ ОТ ВНЕКЛЕТОЧНОЙ КОНЦЕНТРАЦИИ КАЛЬЦИЯ (рисунок из лекций)
Все три стороны процесса возбуждения – электрическая, химическая и структурная – необходимы для понимания его сущности. Но все-таки все начинается с работы ионных каналов. Именно состояние ионных каналов предопределяет поведение ионов, а поведение ионов в свою очередь сопровождается электрическими явлениями. Начинают процесс возбуждения натриевые каналы.
На молекулярно-структурном уровне происходит открытие мембранных натриевых каналов. Сначала этот процесс идет пропорционально силе внешнего воздействия, а затем становится просто «неудержимым» и массовым. Открытие каналов обеспечивает вход натрия в клетку и вызывает деполяризацию. Затем, примерно через 2-5 миллисекунд, происходит их автоматическое закрытие. Это закрытие каналов резко обрывает движение ионов натрия внутрь клетки, и, следовательно, обрывает нарастание электрического потенциала. Рост потенциала прекращается, и на графике мы видим спайк. Это вершина кривой на графике, дальше процесс пойдет уже в обратном направлении. Конечно, очень интересно разобраться в том, что натриевые каналы имеют двое ворот, и открываются они активационными воротами, а закрываются инактивационными, но это следует обсуждать ранее, в теме «Возбуждение». Мы на этом останавливаться не будем.
Параллельно в открытием натриевых каналов с небольшим отставанием во времени идет нарастающее открытие калиевых каналов. Они медлительные по сравнению с натриевыми. Открытие дополнительных калиевых каналов усиливает выход положительных ионов калия из клетки. Выход калия противодействует «натриевой» деполяризации и вызывает восстановление полярности (восстановление электроотрицательности). Но натриевые каналы опережают калиевые, они срабатывают примерно в 10 раз быстрее. Поэтому входящий поток положительных ионов натрия в клетку опережает компенсирующий выход ионов калия. И поэтому деполяризация развивается опережающими темпами по сравнению с противодействующей ей поляризацией, вызванной утечкой ионов калия. Вот почему, пока натриевые каналы не закроются, восстановление поляризации не начнется.
10)
Типы нервных волокон.
Дж. Эрлангер и X. Гассер (1937) впервые классифицировали нервные волокна пс скорости проведения возбуждения.
Классификацию нервных волокон по Эрлангеру - Гассеру давала нам Обухова, в лекции «Функциональная анатомия нервной системы», смотрите там, в учебнике по физиологии есть небольшие расхождения.
В зависимости от скорости проведения возбуждения нервные волокна делят на три типа: А, В, С.
В свою очередь волокна типа А подразделяют на четыре группы: Аα, Aβ, Aγ, Aδ.
Волокна типа А – толстые миелиновые нервные волокна. Эти волокна проводят возбуждение от спинальных моторных нервных центров (двигательных центров спинного мозга) к скелетным мышцам (двигательные волокна) и от рецепторов мышц к соответствующим нервным центрам.
Другие группы волокон типа А (β, γ, δ) имеют меньший диаметр. Волокна этих групп преимущественно проводят возбуждение от различных рецепторов (тактильных, температурных, болевых; рецепторов внутренних органов или висцерорецепторов) в ЦНС, за исключением γ-волокон, значительная часть которых проводит возбуждение от спинного мозга к интрафузальным мышечным волокнам.
К волокнам типа В относятся миелинизированные преганглионарные волокна вегетативной нервной системы.
К волокнам типа С относятся безмиелиновые нервные волокна. Большинство волокон типа С — это постганглионарные волокна симпатического отдела вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления.
Дата: 2019-12-10, просмотров: 694.