Среди мохообразных двудомность распространена шире, чем в других отделах растительного мира [Бойко, 1999]. В мире среди листостебельных мхов двудомными являются более половины видов, среди печеночников – две трети [Longton, Schuster, 1983; Crum, 2001]. В России на долю двудомных листостебельных мхов приходится примерно 54%, причем в различных регионах соотношение одно- и двудомных видов мхов примерно одинаково [Игнатов и др., 2009].
На уровне союзов, для всех типов обследованных лесных сообществ РБ характерно численное превосходство двудомных видов мохообразных над однодомными (рисунок 3).
Установлено, что виды из разных субстратных групп имеют существенные различия в спектрах половых типов видов. В союзах доля однодомных видов варьировала среди эпифитов от 43 до 63 %, среди эпиксилов – от 59 до 87 %, среди эпигейных видов – от 14 до 32 %, среди эпилитных – от 17 до 41 %. Таким образом, однодомные виды преобладали среди видов эпифитно-эпиксильного комплекса, а двудомные – среди напочвенных и эпилитных мохообразных.
Рисунок 3 – Соотношение видов с разными половыми типами в лесных союзах РБ
 |
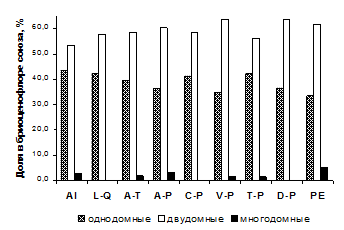
Только для 7 из 42 ассоциаций лесной растительности была выявлена высокая (более 50 %) доля однодомных видов. Такая особенность характерна для сообществ, встречающихся узкими полосами вдоль рек (пойменные ольхово-черемуховые уремники, долинные сосново-березовые леса с периодическим подтоплением), дубняков лесостепной зоны и некоторых типов светлохвойно-широколиственных лесов, которые встречаются небольшими фрагментами среди массивов широколиственных лесов или степей, часто – на крутых склонах. Для всех этих сообществ характерна малоспецифичная бриофлора, представленная, главным образом, ксеромезофитными эпиризными и эпиксильными видами со стратегией колонистов. Развитию двудомных напочвенных мхов в этих сообществах препятствуют такие факторы, как сомкнутый травяной ярус, опад деревьев широколиственных пород, высокий уровень аэрации (в лесостепных сообществах) или поемный режим (в уремниках).
Лесные сообщества РБ часто имеют сложную структуру, встречаются в условиях горного рельефа, в местообитаниях с многочисленными выходами камней. Поэтому использование такого критерия, как соотношение долей однодомных и двудомных видов мохообразных (без учета их субстратной приуроченности) может оказаться недостаточно информативным для того, чтобы делать выводы об экологическом режиме сообществ.
Автор полагает, что для этой цели более оправдано учитывать виды с разными способами размножения. Исследования популяционной биологии мохообразных показали, что расселение за счет спор жизненно необходимо для возникновения новых популяций (при заселении свежих, недавно возникших субстратов, восстановительных сукцессиях, инвазиях в новые растительные сообщества). Вегетативное размножение часто играет ведущую роль для выживания и разрастания популяции, так как оно более эффективно в стабильных условиях, когда происходит клонирование набора генов тех растений, которые оказались хорошо приспособлены к данным конкретным условиям существования [Kimmerer, 2003; Longton, 2006].
Рисунок 4 – Соотношение видов с разными типами размножения среди мохообразных разных субстратных групп
|
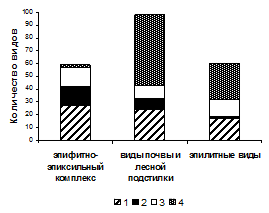
На рисунке 4 показано, что виды групп 1 и 2 преобладают среди эпифитов и эпиксилов. В то же время, почва лесных сообществ и, особенно, каменистые субстраты представляют собой стабильные условия существования, к которым хорошо приспособлены виды мохообразных с преимущественно вегетативным способом размножения, а также виды, не имеющие специализированных органов вегетативного размножения и редкими (или отсутствующим в регионе) спороношением (группы 3 и 4).
Повышенные пропорции видов с активным спороношением отмечены в сообществах союзов Alnion incanae, Lathyro - Quercion, Trollio - Pinion и Dicrano - Pinion. Виды, не имеющие специализированных органов вегетативного размножения и редкое (или отсутствующее в регионе) спороношение, в лучшей степени представлены в сообществах союза Piceion excelsae, в меньшей степени – в сообществах союзов Lathyro - Quercion и Aconito - Tilion.
На уровне бриоценофлор ассоциаций был проведен подсчет соотношения числа видов, часто образующих спорогоны, к числу видов с редким (или отсутствующим в регионе) спороношением. Значение коэффициента меньше единицы оказалось у ассоциаций старовозрастных лесов (Anemonastro biarmiensis - Laricetum sukaczewii , Asaro europaei - Piceetum obovatae , Zigadeno sibirici - Pinetum sylvestris , Cerastio pauciflori - Piceetum obovatae , Linnaeo borealis - Piceetum abietis). Значение коэффициента свыше 1,5 оказалось у лесов, которые либо представлены изолированными участками в поймах (Alnetum incanae), лесостепной зоне ( Brachypodio pinnati - Quercetum roboris ), среди сельхозугодий (Lasero trilobi - Quercetum roboris), либо имеют высокую интенсивность естественных нарушений (Carici arnellii - Pinetum sylvestris , Galio odorati - Pinetum sylvestris).
Таким образом, этот коэффициент может использоваться для оценки стабильности экологических условий в лесном сообществе и косвенно позволяет судить о длительности ненарушенного существования местообитаний. Такие данные, наряду с анализом флористического состава, позволяют выявить наиболее ценные для охраны участки лесов и могут быть использованы для мониторинга за их состоянием.
Дата: 2019-07-30, просмотров: 533.