Группа Carinatae (птицы, имеющие «киль» – выделяющийся участок на грудине для крепления мышц) включает современных беззубых птиц (Aves) и таких их близких родственников, имевших зубы, как ихтиорнитины – «рыбы-птицы». Последние были названы иронично за структуру их позвонков, а не из-за животных, которые почти наверняка составляли основную часть их рациона. Большинство ихтиорнитинов известны по очень фрагментарным остаткам, и, несмотря на то, что некоторые исследователи определили все известные остатки в один единственный вид, их географическое распространение и большой временной промежуток в сочетании с разнообразием размеров во времени, указывают, что некоторые виды потенциально могут быть признаны. Вместе с водными гесперорнитинами килегрудые образуют Ornithurae – продвинутую группу, включающую в себя первых известных по-настоящему водоплавающих птиц. Первой обнаруженной эволюционной линией, специфически адаптировавшейся к нырянию, стали ганьсуиды, известные в настоящее время по единственному виду Gansus yumenensus. Gansus могли быть примитивными килегрудыми, или более базальными эуорнитинами, однако в ряде неофициальных исследований были они были идентифицированы как истинные ихтиорнитины (Cau 2012).
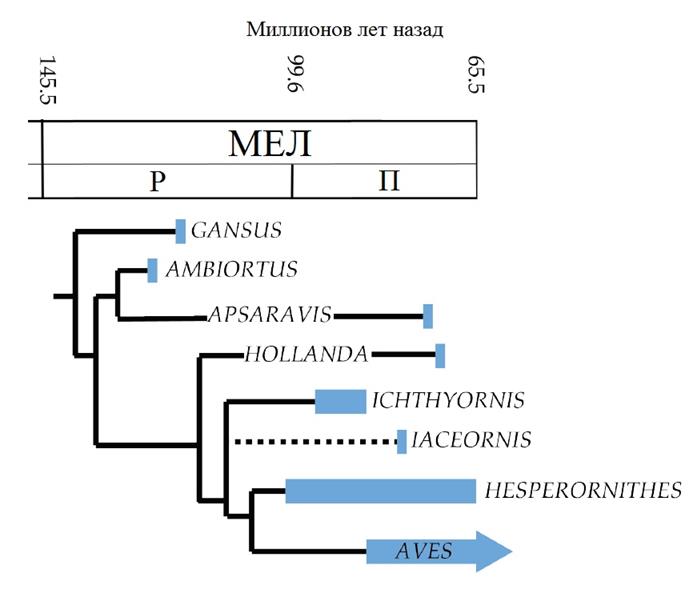
Вверху: Родственные связи килегрудых во временной шкале. Филогения аппроксимирована на основе O’Connor & Zhou 2012.
Колеблющееся начало Деменьтьева Ambiortus dementjevi
Время: 125 млн. лет назад.Место находки: Байконур, Монголия.Среда обитания: формация Андайхудак.Размер: РК неизвестен; ДТ ~25 см (10 дюймов); ПД неизвестна.Характерные особенности: Известен по фрагментарному скелету, отличающемуся внутренней анатомией, включающей тонкие лопатки и расположение креплений связок на плечевой кости. Биология: Длинные, имеющие опахала первичные маховые перья указывают на способность к полёту.В целом очень примитивный, может быть похож на янорнитиформов/предков килегрудых.
Птица Укаан Апсара Apsaravis ukhaana
Время: 75 млн. лет назад.Место находки: Умнеговь, Монголия.Среда обитания: Укаа Толгод, джадохтская свита. Размер: РК >30 см (1 фут); ДТ ~18 см (7 дюймов); ПД неизвестна.Характерные особенности: Голова маленькая, но морда неизвестна.Крылья умеренно длинные (локтевая кость и запястье ~ половина длины туловища) с продвинутым механизмом полёта (заставляющим кисть во время взмаха автоматически раскрываться). Когти крыла отсутствуют. Ноги очень длинные с длинной плюсной (более чем вдвое превышает длину пальца ноги).Пальцы ног относительно толстые с большими, но слабо изогнутыми когтями.Биология: Широкий диапазон движения суставов пальцев ног обычно встречается у видов, приспособленных к бегу или плаванию.Пустынная экосистема, в которой жили представители этого вида, в значительной степени была лишена воды в течение некоторых времён года.В отличие от большинства других мезозойских эуорнитинов, которые в основном занимали морские и прибрежные среды, похоже, что Apsaravis повторно адаптировался к бегу по земле и образу жизни в пустыни.Во время сухих сезонов мог охотится на ящериц и других мелких позвоночных. Возможно, является близким родственником Ambiortus и Palintropus.
Сияющая голландская птица Hollanda luceria
Время: 72 млн. лет назад.Место находки: Умнеговь, Монголия.Среда обитания: формации Хармин-Тсав, Баруун-Гоёот. Пустынное высокогорье. Дюнные поля, склонные к периодическим затоплениям и засушливые лесистые районы.Размер: РК неизвестен; ДТ ~50 см (20 дюймов); ПД неизвестна.Характерные особенности: Известен только по задним конечностям. Ноги очень длинные с длинной, тонкой и сросшейся плюсной. Пальцы стоп сильные и массивные.Биология: Анатомические сравнения ног и пропорций костей пальцев стоп указывают, что представители этого вида кормились на земле, вероятно, преследуя быстро бегающую добычу наподобие современной кукушки-подорожника.Возможно, сохранились зубы в задней части челюстей, как и у большинства других нептичьих эуорнитин.

Юймэнь ГаньсуGansus yumenensis
Время: 120 млн. лет назад.Место находки: Ганьсу, Китай.Среда обитания: формация Сягоу. Широкий бассейн с преобладанием системы больших, спокойных пресноводных озёр. Размер: РК 40 см (1,3 фута); ДТ ~24 см (9 дюймов); ПД неизвестна.Характерные особенности: Крылья большие и мощные, относительно короткие и широкие (плечевая и локтевая кость (каждая из них) составляет приблизительно половину длины туловища, кисть чуть превышает половину длины локтевой кости). Маховые перья асимметричные, первичные маховые перья достигают ~10 см в длину у крупных экземпляров, вторичные маховые до ~8 см. Маленький коготь на пальце крылышка. Малый палец сросся с большим пальцем. Грудина имеет крепкий киль. Ноги длинные (бедренная кость и плюсна приблизительно одинаковой длины, голень почти в два раза больше бедренной кости) с длинными пальцами (равными длине плюсны) и маленькими, прямыми когтями. Стопы перепончатые до основания когтей и покрыты мелкой чешуёй. Биология: Обитали в пресноводных водоёмах. Плавающие и/или ныряющие птицы широко распространены в своей среде. Длинные перепончатые задние конечности приспособлены к гребле по поверхности озера, возможно, и для ныряния за рыбой и беспозвоночными. Вероятно, находясь в воздухе, использовали парящий полёт. Исследования плохо сохранившихся следов перьев у некоторых экземпляров показывают, что оперение было в основном тёмного или чёрного цвета.
Двуглавая рыба-птицаIchthyornis anceps
Время: 80 млн. лет назад.Место находки: Канзас, Алабама, Техас и Калифорния, США; Манитоба, Канада.Среда обитания: стратон Смоки Хилл Чок, формация Ниобара. Глубокие воды западного внутреннего моря.Размер: РК >60 см (2 фута); ДТ ~30 см (1 фут); ПД неизвестна Характерные особенности: Голова большая с длинными, прямыми челюстями, заканчивающимися клювом, поддерживаемым предзубной костью в нижней челюсти. Многочисленные зубы размещены в середине верхней и нижней челюсти сзади от беззубых кончиков. Шея длинная. Крылья длинные и массивные (локтевая кость немного более ¾ длины туловища, плечевая кость и кисть почти равны в длину локтевой кости). Палец крылышка очень короткий и неподвижный. Малый палец сросся с большим пальцем. На всех пальцах крыла отсутствуют когти, но, вероятно, сохранились кератиновые чехлы, по крайней мере на крылышковом и большом пальце, как у настоящих птиц. Грудина имеет крепкий киль. Ноги очень короткие (бедренная кость и плюсна одинаковой длины, голень чуть меньше половины длины бедренной кости, голень немного короче плечевой кости) и очень тонкие. Пальцы стоп в большей степени неизвестны. Биология: Морской вид. Очень распространён вдоль побережья западного внутреннего моря от Альберты до Алабамы. Скорее всего, в экологии подобен чайкам, использовал неглубокие ныряния для ловли рыбы под поверхностью воды. Стопы могли быть перепончатыми. Маленький клюв мог помогать в поимке или пронзании рыбы. Крылья, вероятно, длинные, заострённые и узкие, что позволяло совершать длительные периоды динамического парения. Несмотря на то, что считается мелким видом, известные экземпляры представляют огромный диапазон размеров, а самые крупные были размером с самых крупных чаек, или ещё больше. Вариация размеров не попадает в отдельные классы, как этого следовало бы ожидать, если бы она была представлена несколькими видами, а вместо этого формирует континуум. Вероятно, представляет собой этапы роста одного вида. Это указывает на то, что даже самые продвинутые нептичьи килегрудые росли относительно медленней по сравнению с современными птицами.
Забытая птица Марша Iaceornis marshi
Время: 80 млн. лет назад.Место находки: Канзас, США.Среда обитания: стратон Смоки Хилл Чок, формация Ниобара.Размер: РК неизвестен; ДТ ~25 см (10 дюймов); ПД неизвестна.Характерные особенности: Общей анатомией похож на I. anceps, но внутренней анатомией отличается особенностями строения лопатки и вилочки. Грудина имеет крепкий киль. Ноги гораздо больше по сравнению с крыльями, чем у I. anceps (голень длинней кисти). По крайней мере, на больших пальцах отсутствуют костистые когти. Малый палец полностью сросся с большим пальцем. Биология: Несмотря на то, что очень похож на современный ему вид I. anceps, вероятно, ближе к представителям класса Aves. Неизвестно, имели ли представители этого вида зубы; зубы, скорее всего, отсутствовали, если они относились к настоящим птицам, а их более продвинутое положение указывает на то, что зубы, вероятно, были редуцированы.
Настоящие птицы
Настоящие или «современные» птицы являются ‘вершинной группой’ птиц, потомками самого последнего общего предка всех птиц, живущих в настоящее время. Похоже, что этот предок существовал ещё 130 млн. лет назад, если такие фрагментарные остатки, как Gallornis, действительно, классифицируются относящимися к настоящим птицам.У всех известных настоящих птиц отсутствуют зубы, а вместо них имеются большие, зачастую полностью сросшиеся клювы.Вероятно, что зубы были полностью потеряны у предшественника общего предка всех групп современных птиц; однако также возможно, что две или более эволюционных линий настоящих птиц утратили зубы независимо друг от друга.
Многие виды настоящих птиц были определены из мезозоя, по большей части, из позднего мела, но они основаны на слишком фрагментарных остатках, чтобы с какой-либо долей уверенности можно бы было реконструировать их прижизненный облик (см. Приложение A). Из-за этой скудости летописи ископаемых возник исторически спорный вопрос, касающийся времени диверсификации современных птиц. Однако, вероятное присутствие многих отрядов современных птиц в мезозое указывает на то, что настоящие птицы начали диверсифицировать во многие формы современных птиц ко времени К-Т (мел-палеогенового) вымирания, которое положило конец мезозойской эры.
На основе предварительных интерпретаций ископаемых остатков, можно предположить, что до К-Т границы существовали представители следующих групп современных птиц: Charadriiformes (болотные ржанки, такие как Cimolopteryx), Anseriformes (включая Vegavis, см. выше), Galliformes (включая Austinornis), Palaeognathae (бескилевые и близкородственные виды, представленные Limenavis и, возможно, некоторыми ранними литорнитидами) и Pelecaniforms (в том числе Torotix и некоторые подобные бакланам птицы).Поганки и подобные пастушковым птицы (возможно, предки кайнозойских форусрацидов) также были известны из позднего мела.По крайней мере, один вид или эволюционная линия, представляющая каждую из этих групп пережили К-Т вымирание и диверсифицировали во все известные группы современных птиц кайнозойской эры. В случае с гусеобразными, по крайней мере, два вида или две эволюционных линии пережили массовое вымирание (пресбиорнитиды и предки современных уток и гусей, представленные Vegavis).
Вдобавок к этим эволюционным линиям настоящих птиц, возможно, что одна или более групп, не относящихся к современным птицам, также пережила К-Т вымирание.Один вид из палеогена Qinornis paleocenica известен по окаменелостям, которые демонстрируют несросшиеся кости плюсны, несмотря на определение экземпляра как взрослого, этот признак известен только для эволюционных линий птиц, не относящихся к современным птицам.Этот вид может быть представителем линии зубастых килегрудых птиц, которые прожили в течение нескольких миллионов лет в кайнозое.
Сюда не включена разнообразная орнитофауна формации Хорнерстаун, Нью-Джерси, которая, вероятно, формирует очень короткий промежуток после К-Т границы (о чём свидетельствуют обильные перезахоронения остатков мозазавров). Эта экосистема сохранила обилие водоплавающих птиц, включая болотных птиц и представителей большинства птичьих групп, упомянутых выше. Это ещё одно свидетельство того, что птицы, пережившие вымирание, были преимущественно водоплавающими, позднее они эволюционировали в «высших наземных птиц» и заполнили освободившиеся ниши, оставленные энанциорнитинами и другими тероподами.

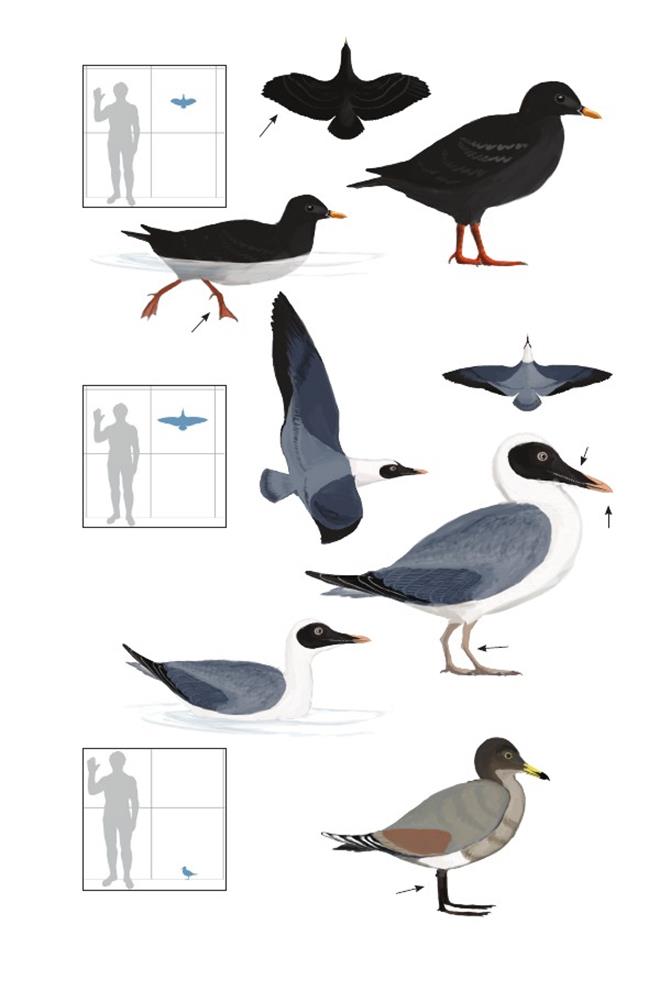
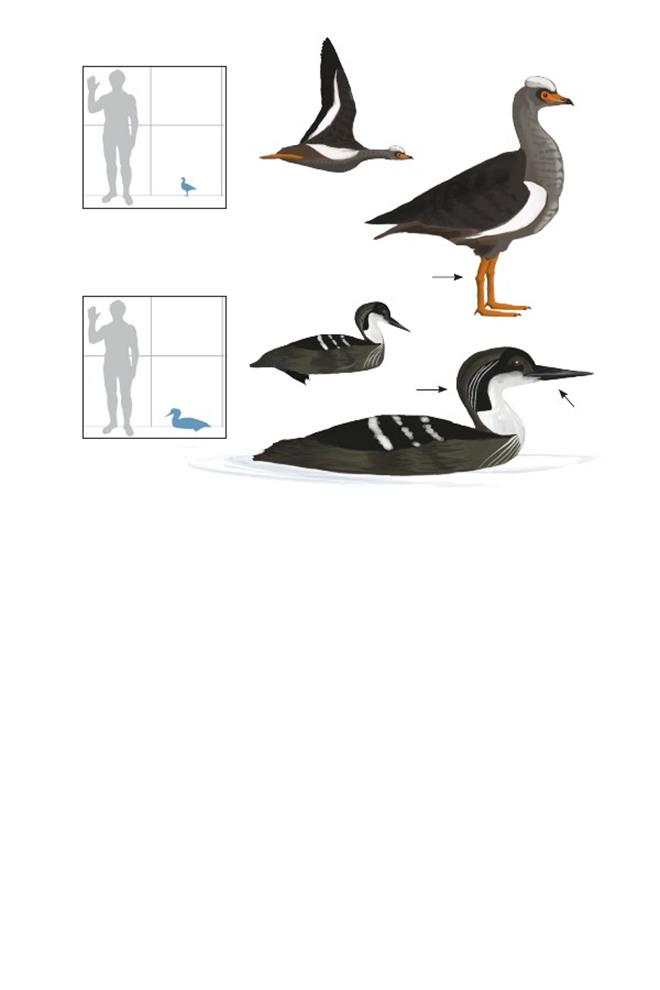
Вверху: Гипотетическая реконструкция отдельных мезозойских представителей настоящих птиц. По часовой стрелке от верхнего левого края: Austinornis lentus, Cimolopteryx rara, Limenavis patagonica, Torotix clemensi. Масштаб не соблюдён.
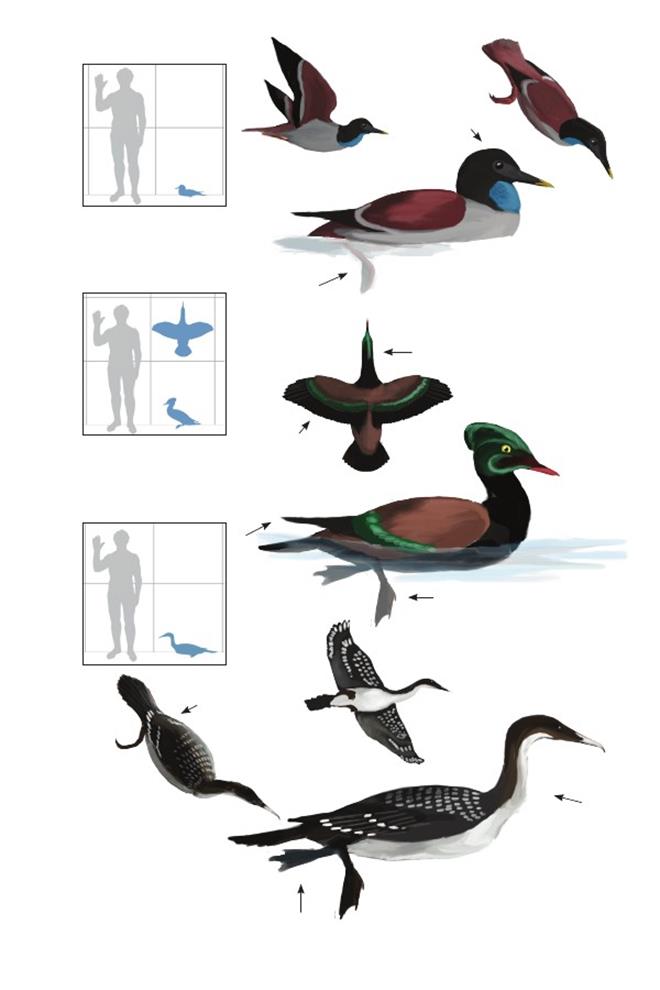
Птица с Веги IAA (Instituto Antartico Argentino) Антарктического аргентинского института Vegavis iaai
Время: 65,5 млн. лет назад.Место находки: остров Вега, Антарктика.Среда обитания: стратон Сэндвич Блафф, формация Лопес-де-Бертодано.Размер: РК неизвестен; ДТ ~25 см (10 дюймов); ПД неизвестна.Характерные особенности: Известен по фрагментарному скелету, серьёзно повреждённому во время препарации из скалистой материнской породы. Крыло известно не полностью.Ноги относительно длинные,голень по длине равняется или превышает длину плечевой кости. Плюсна составляет ¼ или ½ длины локтевой кости (на основе длины лучевой кости).Биология: Относительно длинные ноги могут указывать на болотный образ жизни, похожий на таковой у родственных пресбиорнитидов из палеогенового периода; ширококлювые, длинноногие утки более продвинуты, чем V. iaai.Многочисленные тонкие скелетные признаки показывают, что этот вид был примитивным представителем линии уток, гусей и лебедей (Anatoidea), возможно, похож на таких современных базальных утиных, как паламедеев (Anhimidae) и полулапчатых гусей (Anseranatidae), которые дивергировали от линии уток до того, как обрели характерно широкий, плоский клюв характерный утиным и пресбиорнитидам.Поэтому клюв Vegavis мог быть узким и немного загнутым, как у примитивных гусеобразных, или мог начать расширяться.Это помогло бы точно определить их способ питания, но, как и все базальные гусеобразные, представители V. iaai, вероятно, кормились мягким растительным материалом, либо в воде и/или в заболоченных местах.Как и у паламедеев и полулапчатых гусей, их стопы могли быть частично перепончатыми.
Полярная птица Грегори Polarornis gregorii
Время: 65,5 млн. лет назад.Место находки: остров Вега, Антарктика.Среда обитания: стратон Сэндвич Блафф, формация Лопес-де-Бертодано.Размер: РК неизвестен; ДТ ~80 см (2,6 фута); ПД неизвестна.Характерные особенности: Известен по частичному скелету, состоящему из частичного черепа, позвонков и костей конечностей. Беззубый клюв длинный, узкий и треугольной формы. Голова длинная и низкая.Шея длинная (в полностью вытянутом состоянии почти равна длине туловища).Ноги, вероятно, очень длинные с очень короткой бедренной костью (меньше половины длины черепа) и с большой голенью с участками крепления сильных мышц. Голень пропорционально длинная, но большая её часть неизвестна.Биология: Продвинутые признаки черепа и ног указывают на то, что эти птицы, вероятно, были представителями современной эволюционной линии гагар (Gaviiformes).Стенки их костей были относительно толстыми по сравнению с современными гагарами, что указывает на нелетающий или почти нелетающий, ныряющий образ жизни.Следовательно, и крылья могли быть маленькими по сравнению с таковыми у современных гагар.Neogaeornis wetzeli– другой ранний родственник гагар из Чили, может быть старшим синонимом этого вида или безымянного, очевидно летающего вида из Антарктики, который до сих пор не описан.
Гесперорнитины
Учитывая тот факт, что большинство примитивных эуорнитинов заняло ниши полуводных береговых птиц, не удивительно, что некоторые группы диверсифицировали в полностью водные формы. Hesperornithes или «западные птицы» представляли собой крупную эволюционную линию водных мезозойских птиц. Клада Hesperornithes включает в себя четыре большие линии гребущих ногами ныряльщиков: эналиорнитиды, бродавиды, бапторнитиды и гесперорнитиды. Эналиорнитиды – плохо изученная группа примитивных ныряльщиков, которая может оказаться неестественной группой, включающей предков гесперорнитидов и бапторнитидов. У них, как и у броадависов, вероятно, сохранялась некоторая способность к полёту; перья гесперорнитин, сохранившиеся в янтаре Альберты, возможно, принадлежали гнездившимся эналиорнитидам или бродавидам.
Гесперорнитоиды, включающие в себя наиболее специализированные семейства бапторнитид и гесперорнитид, являлись, насколько это вообще возможно, полностью водными и предельно приспособленными к жизни в море птицами из всех когда-либо существовавших, некоторые из них почти совсем утратили способность к передвижению по суше, и выходили на берег только во время гнездования. У гесперорнитид и современных гагар очень сходна анатомия задних конечностей и тазобедренного сустава; вероятно, при преследовании добычи под водой гесперорнитиды так же гребли лапами. По своему строению пальцы стоп, особенно суставы пальцев стоп, очень похожи на пальцы современных поганок, снабжённые отдельным «лопастями», а не единой перепонкой. Характерной особенностью, вероятно, являлось наличие на одном пальце единой большой лопасти, а не нескольких лопастей, как у лысух. Как и на аэродинамических маховых перьях, лопасти на пальцах, вероятно, также были асимметричными, что улучшало их гидродинамические свойства. Как и ряд специализированных птиц-ныряльщиков, многие из гесперорнитин могли иметь обратно-теневую окраску – тёмную сверху и светлую снизу, делавшую их менее заметными на поверхности и из-под воды. Расположенные в ячейках зубы гесперорнитоидов (такие же зубы наблюдаются и у мозазавров), возможно, были специализированы для захвата быстро двигающейся добычи в открытом океане. Кроме того, как оказалось, они имели челюсти прокинетического типа, то есть верхняя челюсть двигалась вверх и вниз относительно основания черепа, как и у многих современных птиц (Bhuler & al. 1987).
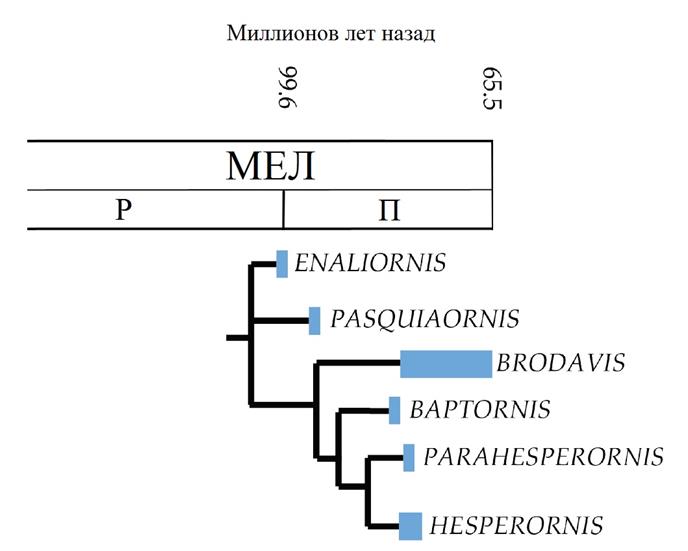
Вверху: Родственные связи гесперорнитинов во временной шкале. Филогения аппроксимирована на основеMortimer 2010.
Морская птица Барретта Enaliornis barretti
Время: 100 млн. лет назад.Место находки: Англия.Среда обитания: формация Кэмбридж Гринсэнд. Берега и лагуны, окружавшие субтропические острова мелководного моря. Размеры: РК неизвестен; ДТ ~55 см (1,8 фута); ПД неизвестна. Характерные особенности: Морские гесперорнитины, известные по частичному скелету и нескольким фрагментам. Голова маленькая, но строение большей части черепа и морды остаётся неизвестным. Ноги очень длинные с длинной мощной голенью, короткой и толстой бедренной костью (вероятно, менее половины длины голени) и короткой, но мощной плюсной, длиною короче бедра. Отличаются малыми размерами, примитивным строением и особенностями анатомии таза и задних конечностей (отсутствие шейки бедра, средний размер антитрохантера, наличие киля под сложным крестцом).Биология:Эти мелкие морские птицы с одной стороны, обладали чертами более продвинутых гесперорнитин, сохраняя в то же время некоторые примитивные признаки, общие с ихтиорнитинами и другими килегрудыми птицами. Следовательно, они, вероятно, представляли собой обитавших на побережье ныряльщиков, и, возможно, сохраняли некоторую способность к полёту, хотя о строении их крыльев ничего не известно.
Птица Танке из Паскиа Pasquiaornis tankei
Время: 95 млн. лет назад.Место находки: Саскачеван, Канада.Среда обитания: формация Эшвилл, прибрежные районы северной части западного внутреннего моря. Размеры: РК >36 см (1,2 фута); ДТ ~70 см (2,3 фута); ПД неизвестна. Характерные особенности: Челюсти длинные, снабжены зубами. Крылья относительно длинные. Ноги длинные и тонкие.Биология:Ноги располагались в большей степени под телом по сравнению с более поздними гесперорнитинами, что, вероятно, способствовало походке вперевалку. Из-за своих пропорций пальцы стоп не могли сгибаться, что указывает на наличие между пальцами единой перепонки, а не отдельных «лопастей». Относительно длинные и массивные кости крыльев указывают на то, что, возможно, эти птицы могли летать, хотя строение грудины неизвестно. Вероятно, это были водоплавающие птицы (Sanchez 2012).
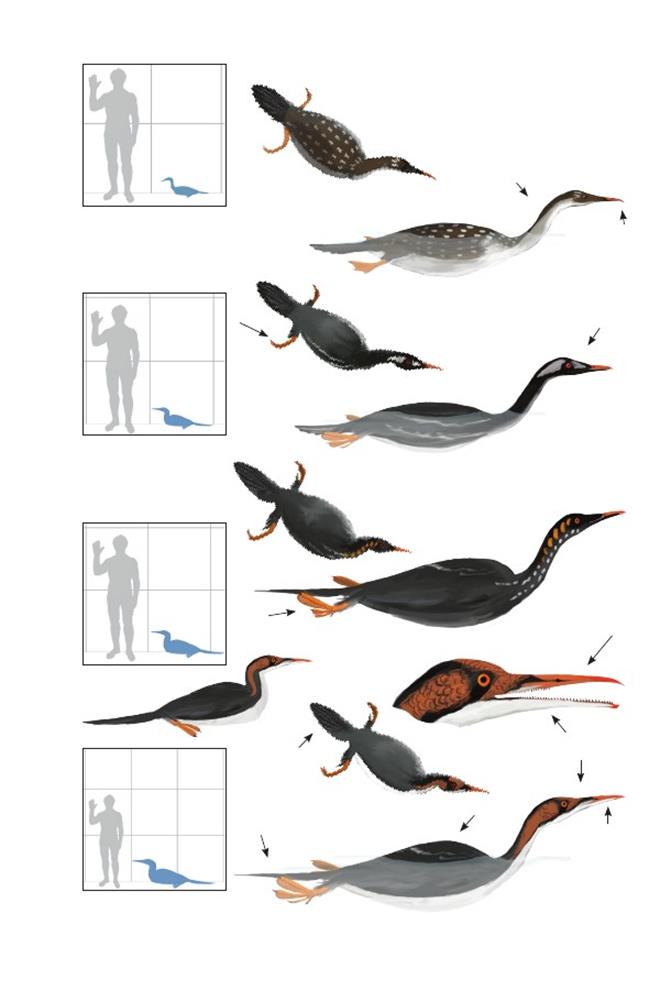
Бродкорбова птица Варнера Brodavis varneri
Время: 78 млн. лет назад.Место находки: Канзас, США.Среда обитания: стратон Шэрон Спрингс, формация Пьер Шейл. Относительно мелкие дельты и устья рек, впадавших во Внутреннее Западное море. Размеры: РК неизвестен; ДТ ~90 см (3 фута); ПД неизвестна. Характерные особенности: Морские гесперорнитины. Тело длинное, бочкообразное, с коротким туловищем и длинными бёдрами. Ноги очень длинные, длина голени превышает половину суммарной длины туловища и бёдер. Плюсна короткая и толстая, менее ¼ длины голени. Шея длинная, по длине, вероятно, равна сумме длин туловища и бёдер. Строение крыльев неизвестно.Биология:Кости таза этих птиц более примитивны и не настолько плотно сросшиеся по сравнению с близкими видами. В сочетании с массивными костями стоп это может указывать на несколько отличающийся способ движения под водой по сравнению с близкими видами, обитавшими в более глубоких морских водах вдали от берегов. В общем, скелет был более лёгким и не таким массивным, как у других гесперорнитин. Это ограничивало способность нырять, что вынуждало этих птиц кормиться у поверхности воды. Более лёгкий скелет может также указывать на сохранение некоторой способности к полёту.
Падающая ныряющая птица Baptornis advenus
Время: 80 млн. лет назад.Место находки: Канзас, США.Среда обитания: стратон Смоки Хилл Чок, формация Ниобара. Глубокие воды тёплого внутреннего моря с преобладанием аммонитов и разнообразных рыб – от мелких до гигантских. Размеры: РК 36 см (1,2 фута); ДТ ~70 см (2,3 фута); ПД неизвестна. Характерные особенности: Морская птица. Голова длинная и узкая, очень узкая снабжённая зубами морда оканчивалась клювом умеренной длины. Шея длинная и тонкая. Тело длинное, бочкообразное. Крылья маленькие и рудиментарные. Ноги длинные, но с короткой плюсной и маленькими стопами.Биология:Самый мелкий вид гесперорнитин в этой среде. Вероятно, охотился на более мелкую добычу и в более мелких водах, чем родственные виды. Анатомия суставов пальцев стоп указывает, что, вероятно, пальцы были соединены перепонкой и не имели отдельных «лопастей».
Почти западная птица Алекса Parahesperornis alexi
Время: 80 млн. лет назад.Место находки: Канзас, США.Среда обитания: стратон Смоки Хилл Чок, формация Ниобара.Размеры: РК неизвестен; ДТ 1,1 м (3,6 фута); ПД неизвестна. Характерные особенности: Похож на Hesperornis, но более примитивный. Анатомия пальцев стоп в большей степени характерна для птиц с «лопастями» на пальцах, нежели с перепонками.Биология: Единственные известные остатки гесперорнитин с сохранившимися отпечатками кожи и перьев. Как и у поганок, плюсна была спереди покрыта широкими чешуйками (всего 26 чешуек). Ближе к туловищу, плюсна была покрыта очень длинными пуховыми перьями, концы которых доставали почти до подошвы стопы.
Западная птица с толстой подошвой стопы Hesperornis crassipes
Время: 80 млн. лет назад.Место находки: Канзас, США.Среда обитания: стратон Смоки Хилл Чок, формация Ниобара Размеры: РК неизвестен; ДТ 1,1 м (3,6 футов); ПД неизвестна.Характерные особенности: Морская птица, по своей анатомии близка H. regalis. Отличается строением плюсны с более высокими мышечными бугорками на лодыжке, и менее выраженными местами крепления мышц на грудине.Биология: обитали вместе с родственными видами, вероятно, используя другие экологические ниши. Различия в анатомии плюсны могут указывать на различные способы гребли задними конечностями при нырянии.
Королевская западная птица Hesperornis regalis
Время: 80-78 млн. лет назад.Место находки: Канзас, США.Среда обитания: стратон Смоки Хилл Чок, формация Ниобара Размеры: РК ~46 см (1,5 футов); ДТ 1,7 м (5,5 футов); ПД неизвестна.Характерные особенности: Голова маленькая с длинными тонкими челюстями, оканчивающимися сложным клювом. Позади клюва на челюстях расположены многочисленные зубы. Клюв на нижней челюсти ограничен предзубной костью, на верхней челюсти клюв тянется дальше назад, покрывает длинную предчелюстную кость и продолжается чуть выше над зубами. Клюв немного загнут. Зубы нижней челюсти размещаются в ячейках и при смыкании челюстей замыкались в ямки предчелюстной кости. Тело короткое и округлое с изогнутой спиной. Крылья очень маленькие, вероятно, рудиментарные и скрыты под покровными перьями. Верхняя часть ног крепилась к туловищу, и, вероятно, крепилась к стенке тела мягкой тканью. Плюсны и стопы расходятся в стороны от области таза/хвоста. Пальцы стоп длинные, четвёртый (внутренний) палец самый длинный. Вероятно, пальцы стоп имели отдельные «лопасти». Хвост широкий и плоский, возможно, напоминал хвост бобра.Биология: Известен по многочисленным остаткам, в том числе по почти полным скелетам. Как и многие крупные глубоко ныряющие морские птицы, они могли иметь обратно-теневую маскирующую окраску. Вероятно, во время ныряния шея фиксировалась связками в кривую с резким изгибом, а голова прижималась к туловищу, что позволяло избежать скручивания шеи во время быстрых поворотов под водой. Учитывая, насколько задние конечности были растопырены и прижаты к обтекаемому туловищу, так же, как и у гагар, они, вероятно, были крайне неповоротливы на суше и неспособны ходить, а вместо этого толкали своё тело вперёд и скользили на животе, ползая, наподобие современных тюленей (Martin & al. 2012).
Приложение A: исключённые виды
К сожалению, многие виды ископаемых птиц были выделены лишь по крайне фрагментарным остаткам или же по остаткам, которые невозможно реконструировать, поскольку их классификация остаётся недостаточно определённой, чтобы выполнить реконструкцию, основываясь на реконструкциях родственных видов. Эти виды кратко описаны ниже.
Дата: 2019-07-24, просмотров: 575.