Несмотря на пренебрежение со стороны популярных средств массовой информации, акцентирующих внимание на более крупных чешуйчатых динозаврах, мезозойские птицы были очень разнообразны.Согласно ранним классификациям, этих первых «птиц», в целом, можно разделить на два типа: с длинным и коротким хвостом.Классификации, предшествующие кладистическим, разделили их на подклассы «Sauriurae» («ящерохвостых», получивших своё название за длинные костистые хвосты, похожие на хвосты рептилий) и «Ornithurae» («птицехвостых», у которых развился резко укороченный хвост современного типа).Однако эти подклассы представляют собой эволюционные стадии, а не клады. Короткохвостые птицы произошли от длиннохвостых предков и, таким образом, длиннохвостые птицы в целом не могут считаться отдельной группой.
Chiappe (2006) назвал эти два основных типа «веерохвостыми птицами» и «вайяхвостыми птицами» (от «вайя» – лист папоротника) из-за различного расположения перьев, определяемого разной длиной хвоста. У вайяхвостых птиц (более примитивный тип) хвостовые (рулевые) перья, как правило, располагались хотя бы частично, вдоль костного хвоста, образуя структуру, напоминающую ветку папоротника. Каждая пара рулевых перьев, как правило, крепилась к одному из хвостовых позвонков. Современные птицы являются веерохвостыми. Вследствие уменьшения хвоста, все рулевые перья крепятся к одной лемехообразной кости, называемой пигостилем, и образуют веер, который птица может часто складывать и раскрывать с помощью мышц, управляющих рулевыми перьями.
Однако это различие среди мезозойских птиц не является твердо установленным.Настоящие веерохвостые птицы, возможно, возникли позже, чем полагалось сначала. Многие ранние короткохвостые птицы – такие, как имеющие клюв длиннокрылые конфуциусорнитиды, имели только пару специализированных лентообразных перьев на хвосте, причем только у представителей одного пола. У более половины известных экземпляров Confuciousornis sanctus рулевые перья отсутствовали, а их хвосты представляли собой лишь пушистые бугорки, покрытые контурными перьями. Формы с короткими хвостами могли быть самками, или особями, переживающими период линьки. Лентообразные перья (предположительно) самцов, которые также найдены у многих экземпляров энанциорнисов («противоптиц»), как правило, были парными. Присутствовало два или четыре пера, но известны некоторые виды, у которых их число достигало восьми.У всех у них на кончике хвоста имелись сросшиеся укороченные хвостовые позвонки, напоминавшие пигостиль современных птиц. Однако у тех птиц слитная структура хвоста была простой и имела форму стержня или кинжала, и вовсе не была похожа на сложный пигостиль современных птиц с крепящейся к нему мускулатурой. Таким образом, хвостовое оперение не могло быть раскрыто или сложено для облегчения поворота во время полёта или для приземления. Этих промежуточных короткохвостых птиц за их уникальное строение, пожалуй, лучше было бы назвать «лентохвостые птицы».
Настоящие веерохвостые птицы, заведомо, более прогрессивные, включают в себя, в основном, современных птиц (Aves) и их ближайших эволюционных родственников (вместе их называют эуорнисовыми, «собственно птицами»). Они характеризуются наличием веера из упорядоченных по размеру рулевых перьев, в котором, по крайней мере, два самых задних пера крепятся непосредственно к лемехообразному пигостилю. Пигостиль в свою очередь связан с мышечным органом, называемым bulbi rectricium, к которому крепятся все рулевые перья. Мышца, складывающая и раскрывающая перья наподобие веера, позволяет достичь гораздо большей манёвренности, чем у предковых видов. Без такой структуры хвостовые перья птиц были бы почти неподвижными и в большинстве случаев служили бы только для демонстрации.
Ваяхвостые, лентохвостые и веерохвостые птицы сосуществовали во время быстрой диверсификации авиалов в раннем мелу. Большинство летающих птиц конца мелового периода были представителями двух основных эволюционных линий короткохвостых: Enantiornithes (лентохвостые) и Euornithes (веерохвостые). Enantiornithes были доминирующими летающими птицами в мезозое, однако все они вымерли 66 миллионов лет назад вместе с большинством других сохранявшихся эволюционных линий динозавров.Выжило лишь несколько линий эуорнитинов, все они были представителями группы, от которой, в конечном итоге, произошли бескилевые, куроподобные и ржанковые, и которые в течение кайнозойской эры диверсифицировали во все другие современные группы птиц.
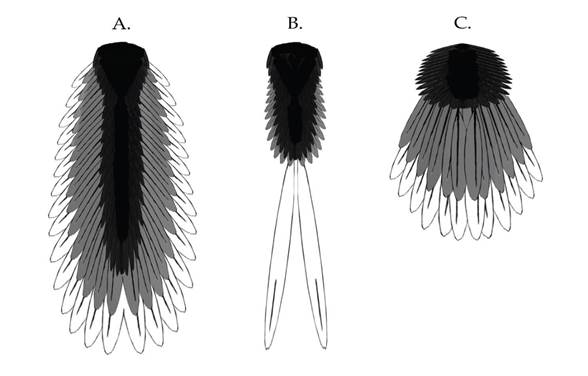
Вверху:Основные типы хвостов мезозойских птиц. A) папоротникообразный или листообразный (например, у базальных авиалов); B) лентообразный (например, у энанциорнисов); C) веерообразный (эурорнисы)
Эволюция полёта
Современным птицам, возможно, в наибольшей степени свойственна их способность к полёту, т.е. создание подъёмной силы собственными усилиями.Большинство современных групп птиц является активно летающими, а в прошлом исследователи часто поддавались соблазну уравнять «птицу» и «полёт».Однако наши современные знания о летописи ископаемых указывают на то, что происхождение птиц (или, по крайней мере, перьев современного типа стадии V) должно было предшествовать возникновению активного полёта.
Изначально возник спор по поводу происхождения полёта: случилось ли это «с земли вверх», т.е. птицы произошли от сугубо наземных предков, или же лазающие по деревьям предки птиц полетели «с деревьев вниз». Но подобное противопоставление представляется ошибочным, так как вполне возможно, что в итоге к возникновению полёта у современных птиц привела некая комбинация из двух возможных способов взлёта. Тем не менее, обе гипотезы, действительно, делают эти предположения проверяемыми, т.е. их можно сравнить с доступной (правда, ограниченной) летописью окаменелостей. Происхождение полёта строго «с деревьев вниз» предполагает, что предки птиц уже были древесными существами, приспособленными для лазания. Согласно этой модели также было бы вероятно, что развитие полёта у животных, использующих свои крылья для затяжных прыжков с пассивным скольжением между ветками деревьев, изначально проходило через фазу планирования.
С другой стороны, модель строго «с земли вверх» предполагает изначальное развитие машущего полёта, так как бегающие по земле животные использовали свои крылья для совершения затяжных прыжков, и, чтобы в конечном итоге оторваться от земли, отталкивались от крутых склонов. К тому же это может означать, что обращённый назад большой палец стопы начал развиваться только после окончательного обретения способности к взрывному взлёту, позволив ранее наземным животным садиться на ветви деревьев (Ohmes 2012).
К сожалению, с помощью летописи окаменелостей не удалось убедительно подтвердить ни один из этих сценариев. Самые примитивные крылатые динозавры, такие как Microraptor и Confuciusornis, имели задние конечности явно лишённые каких-либо выраженных приспособлений для лазания.У предшественников птицегрудых хватающая стопа была ещё несовершенной, тогда как активный полёт, вероятно, уже был освоен, но первые этапы развития большого пальца стопы заметны уже у конфуциусорнитидов, если не у дейнонихозавров.Однако анатомия плеча этих примитивных форм тоже не позволила бы им совершать взрывной взлёт, и, вероятно, они были неспособны взлетать с земли. Возможно, даже ранние птицегрудые, чтобы подняться в воздух, были вынуждены карабкаться на дерево.
На данный момент единственной убедительной гипотезой о происхождении полёта является промежуточная гипотеза. Вполне вероятно, что у предков птиц полёт и древесный образ жизни на самом деле коэволюционировали. Очевидно, что древесные адаптации предков птиц эволюционировали одновременно с приспособлением к полёту.Вероятно, что самый рудиментарный планирующий полёт или бег по наклонной поверхности практиковались самыми ранними крылатыми птицами, и ареал обитания этих животных постепенно расширялся на деревья, где они могли спасаться от хищников или охотится вне досягаемости своих конкурентов. Это можно рассматривать как гибридную гипотезу «с земли – верх на деревья». Как только птицы стали полностью древесными обитателями, полёт усовершенствовался, но на начальных этапах развивался параллельно с адаптацией к древесному образу жизни.
У современных птиц полёт приобрёл различные формы, которые в целом можно охарактеризовать пятью различными стилями: машущий, машуще-скользящий, волнообразный, парящий и взрывной (Close & Rayfield 2012). Машущий полёт представляет собою непрерывные взмахи и используется такими современными птицами, как утки и фламинго; во время полёта они почти непрерывно совершают взмахи.Птицы, использующие машущий полёт, имеют разные размеры, но их крылья, как правило, испытывают высокие нагрузки (т.е. небольшие размеры крыльев по отношению к массе тела птицы).Машуще-скользящий полёт является распространённой формой полёта, он наблюдается у ворон, чаек и многих других птиц, машущий полёт у них непостоянный и периодически сменяется раскрыванием крыльев для скольжения по воздуху.Волнообразный полёт является ещё одной формой прерывистого полёта, наблюдаемой в основном у мелких птиц, где вместо скольжения птица между взмахами крыльев входит в баллистическую фазу. Крылья в этот момент сложены вдоль тела, а не раскрыты, в результате чего тело птицы приобретает в воздухе улучшенную аэродинамику и напоминает пулю.Парящий полёт включает в себя две подкатегории полёта: динамическую и статическую. При статическом парящем полёте нагрузка на крыло невелика, такие птицы имеют широкие крылья с щелевидными просветами на концах, что позволяет им пассивно использовать энергию восходящих тепловых потоков.Статически парящие птицы в основном представлены такими крупными видами, как стервятники.Динамически парящие птицы используют энергию волн воздуха, обычно вблизи поверхности воды, и, как правило, имеют длинные узкие крылья и испытывают более высокие нагрузки на крыло. К ним относятся такие большие морские птицы, как альбатросы и бакланы. Взрывной полёт встречается, в основном, у наземных птиц, способных летать только на очень короткие расстояния, поскольку при полёте они не могут долго махать крыльями, к ним относятся, например, перепела. Эти птицы, как правило, используют полёт для того, чтобы спастись от хищников (Close & Rayfield 2012).
Исследование Close & Rayfield (2012) обнаружило грубую корреляцию между этими пятью стилями полёта и формой вилочковой кости, позволяющую сделать вывод о стилях полёта некоторых мезозойских птиц. Как и следовало ожидать, самые примитивные птицы, такие, какArchaeopteryx и Confuciusornis, у которых были широкие U-образные вилочки, оказались в группе с нелетающими, парящими и планирующими птицами. Впрочем, вероятно, это связано с тем, что у этих примитивных птиц ещё не могли развиться вилочки прогрессивной формы. Это подтверждают и другие исследования, указывающие, что такие птицы были способны совершать только слабые махи, если они вообще имели место, и, вероятно, полеты были преимущественно планирующими.
Как и следовало ожидать, из-за большого разнообразия энанциорнисов, они, вероятно, использовали широкий спектр стилей полёта, в том числе машуще-скользящим и волнообразным. Тем не менее, согласно проведённым в 2012 году исследованиям, многие энанциорнитины не входят ни в одну группу современных птиц.Это связано с уникальным строением их вилочек, имевшим V-образную форму и длинный выступ назад, который частично мог взять на себя роль киля грудины эуорнитинов.Таким образом, энанциорнитины, возможно, не освоили стандартные стили полёта из-за того, что задействовали другие группы мышц, нежели современные птицы, или же они обладали уникальным стилем полёта, исчезнувшим к настоящему времени (Close & Rayfield 2012).
Подобного рода исследования предполагают, что полёт появился у самых примитивных птиц, но не «усовершенствовался» (т.е. не достиг стиля полёта современных птиц), пока в раннем мелу не появились энанциорнитины.Вероятно, в отличие от современных птиц, неэуорнитиновые птицы не умели столь хорошо летать (или, по крайней мере, обладали очень разными видами полёта). Нептицегрудые птицы (более примитивные, чем энанциорнитины) не были способны совершать полный взмах из-за сравнительно близкого расположения плечевого пояса к животу, а не к боковой или задней части грудной клетки. Это позволяло лишь скользить по воздуху или же, в лучшем случае, совершать при полёте только короткие и слабые взмахи крыльями. Энанциорнитины, скорее всего, приобрели способность к полному взмаху, но, вероятно, ещё не обладали такими движениями крыльев вниз, как у современных птиц, когда передняя конечность движется во внешнюю сторону и вперёд (разгибаясь) в результате чего активный полёт становится менее энергозатратным при создании подъёмной силы (Close & Rayfield 2012).Как обсуждалось выше, у энанциорнитинов сократились хвосты и, соответственно, исчезла большая часть хвостовых перьев.Это серьёзно ограничивало их способность контролировать свою скорость полёта, а точное приземление становилось практически невозможным. Наличие удерживающих когтей на крыльях у этих видов указывают на то, что при движении среди деревьев, взлёте и приземлении они в большой степени всё ещё могли полагаться на лазание.
Развитие у энанциорнисов веерообразного хвоста и рулевых перьев способствовало повышению манёвренности и управления скоростью, хотя у многих примитивных видов веерообразный хвост был длинный и экстравагантный; это указывает на то, что изначально такой хвост мог развиться для демонстрационного поведения, возможно, являясь усложнённым лентообразным хвостом более примитивных птиц. Вскоре у эуорнитин когти крыльев редуцировались, что указывает на исчезновение потребности лазать. В значительной степени это может быть обусловлено сочетанием наличия веерообразного хвоста и переходом к водному образу жизни, наподобие прибрежных птиц, а также более совершенной анатомией крыла, что расширило возможности взлёта с ровной поверхности или с поверхности воды.
Реконструкция мезозойских
Птиц
Поскольку мезозойские птицы и другие ископаемые динозавры давно вымерли, проиллюстрировать их можно лишь в разумных пределах погрешности, используя анализ ископаемых свидетельств в сочетании с принципами, почерпнутыми из наблюдений за современными видами и за подходящими ныне живущими аналогами. Благодаря небольшому, но всё увеличивающемуся числу остатков ископаемых птиц с сохранившимися кожей и перьями, можно получить общее представление о возникновении перьев, их распространённости среди первых птиц и их предков, а также о различиях типов и форм перьев у основных групп животных.
Тем не менее, следует иметь в виду, что для всестороннего изображения доисторических птиц приходится допускать множество предположений.Почти все признаки, по которым мы отличаем один ископаемый вид от другого, являются скелетными, и большинство из этих различий при жизни птицы были скрыты перьями и другими мягкими тканями.Разные виды птиц почти всегда заметно различаются расцветкой, гребнями, окрасом или поведением, в общем, теми особенностями, которые в ископаемом состоянии сохраняются крайне редко.
Представьте себе, что путешествующая во времени палеонтолог попадает в джудитский век канадского штата Альберты, 75 миллионов лет назад. Она наблюдает в естественной среде обитания два похожих, хотя и очевидно разных вида длиннохвостых наземных птиц, отличающихся лишь поверхностно: своим окрасом.Ей известно, что в это время в этом месте жило шесть видов длиннохвостых наземных птиц: ценагнатиформы Chirostenotes pergracilis и безымянный вид авимимид, эудромеозавры Dromaeosaurus albertensis, Hesperonychus elizabethae,Saurornitholestes explanatus и троодонтиды Troodon formosus. Некоторых из них легко исключить: прежде всего, это ценагнатиформы с их маленькими головами, клювами, толстыми хвостами, а также не имеющие втягивающихся когтей на вторых пальцах стоп.D. albertensis, вероятно, можно отличить по их высокой, квадратной морде. H. elizabethae, скорее всего, будет отличаться своими маленькими размерами и большими крыльями и, возможно, наличием ножных крыльев. Однако S. explenatus и T. formosusбыли близки по размерам (особенно молодые особиT. formosus). Стопы и задние конечности S. explanatusбыли шире и массивнее, а морда T. formosusпри взгляде сверху – более острой, но эти детали можно рассмотреть только с близкого расстояния.Троодонтиды могли иметь, а могли и не иметь на хвосте листовидного расширения, более длинного и широкого по сравнению с эудромеозаврами, как правило, такое расширение на кончике хвоста имевшими, но этот признак нельзя использовать для уверенного различения видов без предварительного подтверждения. С целью надежной идентификации этих видов палеонтолог должна поймать и изучить или даже препарировать несколько экземпляров, чтобы сопоставить анатомию скелета с только что обнаруженными ею видимыми различиями цвета оперения.
Этот сценарий демонстрирует ряд проблем, связанных с созданием справочника вымерших животных.Однако значение таких справочников в том, что они иллюстрируют наши общие представления о разнообразии вымерших животных, пусть даже основанные на большом количестве предположений, и при этом указывают на достоверно идентифицированные отличительные черты.В некоторых случаях я преднамеренно чрезмерно широко распространял известные признаки нескольких видов на целые группы. Например, некоторые троодонтиды (Anchiornis huxleyi, Jinfengopteryx elegans) сохраняли листовидное расширение хвостовых перьев, которое охватывало почти весь скелет хвоста до его основания.Но мы не можем быть уверены, что этот признак присущ всем остальным троодонтидам, и, хотя весьма вероятно, что существовали исключения, я реконструировал большинство троодонтидов с длинными хвостовыми расширениями. Точно так же и большинство окаменелостей других известных орнитодесмидов и базальных авиалов имели короткие листовидные расширения, ограниченные на кончике хвоста, и снова для облегчения идентификации я перенёс эту частную особенность на всех эудромеозавров как объединяющий их признак.
О некоторых признаках, таких, как распределение и типы перьев, строение клюва, зубов и даже окрас, можно сделать более уверенный вывод на основе эволюционных взаимосвязей и экологии: этот процесс объясняется ниже в следующих разделах.

Вверху:Troodon formosus (сверху) и Saurornitholestes explanatus (снизу) реконструированы с одинаковой длиной, с идентичным оперением и расцветкой.
Перья и крылья
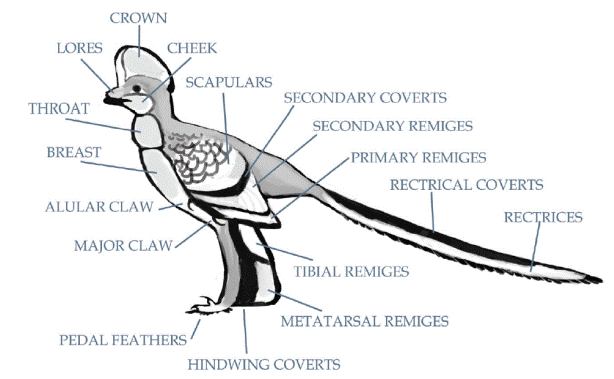
Классификация перьев основывается на их анатомии, расположении на теле и выполняемых функциях. Существует два основных типа перьев. Контурные перья имеют жесткую центральную «нить» или стержень, от которого ответвляется множество мелких волокон, называемых бородками, связанных друг с другом в единое опахало крошечными бородочками и микроскопическими крючочками. Такое расположение бородок, бородочек и крючочков позволяет опахалу разделяться и «застёгиваться» обратно, наподобие липучки. Пуховые перья имеют короткие центральные стержни и бородки без бородочек и крючочков, это не позволяет сформироваться опахалу и приводит к образованию мягких пучков. Пуховые перья, как правило, образуют теплоизолирующий слой и размещаются под контурными перьями птицы. Контурные перья крыла называются маховыми, а перья хвоста – рулевыми. Перья туловища называются покровными, они мягче, и, как правило, расположены более рыхло, хотя по-прежнему относятся к контурным и имеют опахало.
У мезозойских птиц так же, как, в общем, и у современных птиц, перья, покрывающие нижнюю часть головы, зачастую начинались в той же точке (или около неё), что и вышележащие перья, и были очень длинными, что придавало голове птицы характерные округлые очертания. Точно так же большинство пернатых ископаемых птиц обладали очень длинным оперением шеи. Тем не менее, большинство манирапторов давно изображают с жилистыми и стройными шеями, хотя на самом деле она скрывалась в пышном оперении и выглядела довольно короткой, за исключением тех моментов, когда животное вытягивало шею во всю длину.
У большинства современных птиц плюсны и пальцы ног лишены перьев, а верхнюю часть стоп покрывает чешуя. То же самое, как представляется, имело место и у большинства мезозойских птиц, за исключением самых примитивных. Ноги многих самых базальных птиц покрывали длинные перья, на пальцах сменявшиеся более короткими. Они могли быть первыми пернатыми животными, и были полностью покрыты перьями, а чешуя только на ногах повторно развилась позже. С другой стороны, у некоторых видов перьевой покров на задних конечностях мог быть генетическим следствием развития ножных крыльев.
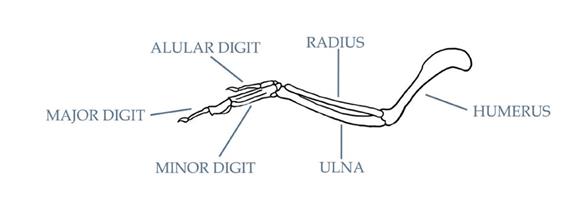
Общей чертой птицеподобных динозавров и ранних птиц были «передние лапы» или «руки». Многие из этих животных имели вполне сформировавшиеся крылья, в которые превратились верхние конечности. Маховые перья покрывали нескольких первых фаланг и пястных костей передних конечностей, представляя собой такие же важные части «рук», как и когти. Связки соединяли перья крыла с мышцами и костями крыла, часто оставляя следы в виде маховых или локтевых бугорков. Эти связки позволяют перьям двигаться или складываться по отношению к костям крыла. Такое движение обычно сопровождается складыванием запястья назад, оно стало возможным благодаря наличию запястной кости в форме полумесяца, называемой полулунной костью запястья. Угол, на который запястье может складываться, у примитивных птиц варьирует, запястье по-настоящему плотноприлегает только у ак-
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Вверху: Иллюстрация Anchiornis huxleyi с маркировкой крупного оперения
тивных летунов.Несмотря на то, что точный угол складывания запястья у ископаемых видов рассчитать трудно, некоторым исследователям удалось выполнить максимально точные оценки, основываясь на таких показателях, как угол лучевой кости в запястье (Sullivan et al. 2010). Самые примитивные паравесы (включая археоптеригидов, базальных троодонтидов, базальных орнитодесмидов и т.д.) могли складывать запястья на угол около 90-100 градусов относительно локтевой кости (Senter 2006). У некоторых нелетающих линий, таких, как эудромеозавриды, этот угол уменьшился. К примеру, Deinonychus мог складывать свои крылья только лишь под углом примерно 120 градусов, вероятно, это обусловлено большей ролью крыла в хищническом поведении (Senter 2006). Угол складывания значительно увеличился у летающих линий, ведущих к современным птицам, и часто достигает 60 градусов между пястью и локтевой костью. Интересно, что, по крайней мере, некоторые ценагнатиформы могли складывать свои запястья даже на бόльший угол, чем большинство современных птиц (Sullivan et al. 2010).
Обычно птицы имеют три пальца на руке (кисти). Первый, самый короткий палец, называется крылышковым (назван так потому, что он крепится к крылышку у современных птиц, это обсуждается ниже). Второй, самый крупный и длинный палец, называется большим пальцем. Третий палец называется малым. Малый палец, обычно длинный и тонкий у примитивных птиц, у многих активных летунов редуцировался, а у современных птиц, как и у некоторых вымерших линий, сросся с большим пальцем. Вполне вероятно, что даже у некоторых более примитивных птиц малый палец был соединён с большим пальцем кожей и мягкими тканями. Когти (когтевые фаланги) представляют собой примитивную черту у птиц – предки птиц имели когти на каждом из трёх пальцев. Сравнительные размеры и изгибы когтей крыльев у ранних птиц существенно различались, они могли использоваться для любых целей, от чистки перьев до лазания, охоты и драки. У более продвинутых птиц некоторые или все когтикрыла утрачены. Подобная редукция когтей крыльев происходила, по крайней мере, в двух отдельных линиях (энанциорнисовой и эуорнисовой), и в обеих группах у видов с увеличением машущей способности. Вполне вероятно, что большинство примитивных авиалов использовало когти крыльев преимущественно для того, чтобы забраться вверх на достаточную высоту и оттуда взлететь. Когда машущий полёт развился до такой степени, что для этих птиц стало возможным отталкивание от земли, роль этих когтей снизилась. Когти могли сохраняться как рудиментарные органы у некоторых линий; фактически же некоторые современные птицы сохраняют чехлы небольших когтей на крылышковых пальцах.
Внешняя часть крыла состоит из первичных перьев, которые крепятся к большому пальцу (к костям пястья и фалангам). Опахала этих перьев у летающих птиц, как правило, очень ассиметричны, ширина опахала с обеих сторон от стержня неодинакова. У многих птиц первичные маховые перья накладываются друг на друга, образуя ровный край крыла, однако у некоторых видов внешние первичные перья разделены и формируют зубчатую линию, что улучшает манёвренность. У некоторых примитивных птиц (кости кисти которых не срослись и не сформировали пряжку) гибкость пальцев крыла могла бы способствовать дополнительной манёвренности за счёт изменения изгиба крыла и расстояния между перьями.
 Вторичные перья, которые крепятся к задней части кости предплечья (к локтевой кости), как правило, шире и более округлые, чем первичные. Вторичные маховые перья обычно полностью формируют внутреннюю часть крыла и направлены немного в сторону тела по мере приближения к локтю. Это придаёт внутренней части крыла округлый профиль и помогает перекрыть любой промежуток, который образуется между крылом и телом из-за коротких или отсутствующих третичных перьев.
Вторичные перья, которые крепятся к задней части кости предплечья (к локтевой кости), как правило, шире и более округлые, чем первичные. Вторичные маховые перья обычно полностью формируют внутреннюю часть крыла и направлены немного в сторону тела по мере приближения к локтю. Это придаёт внутренней части крыла округлый профиль и помогает перекрыть любой промежуток, который образуется между крылом и телом из-за коротких или отсутствующих третичных перьев.






 Перья третьего порядка, или третичные, присутствуют у некоторых птиц, у которых они крепятся к верхней части крыла (к плечевой кости). Обычно большинство птиц держат верхнюю часть крыла в основном прижатой к телу, даже во время движений крылом, и поэтому третичные перья у них либо отсутствуют, либо редуцированы до контурных перьев, подобных тем, которые располагаются на теле. Только специализированные на парящем полёте птицы, которые вытягивают крылья на всю их длину почти в прямую линию, имеют третичные перья, похожие по внешнему виду на вторичные маховые. Лишь у некоторых птиц известны третичные маховые перья, хотя они могли присутствовать у других эуорнитин, таких какIchthyornis, которые, вероятно, были чайкоподобными динамически парящими птицами. У более примитивных птиц, таких, как Archaeopteryx, третичные маховые перья отсутствовали, но имелись перья подобные контурным (а также лопаточные), частично заполнявшие промежуток между крылом и телом.
Перья третьего порядка, или третичные, присутствуют у некоторых птиц, у которых они крепятся к верхней части крыла (к плечевой кости). Обычно большинство птиц держат верхнюю часть крыла в основном прижатой к телу, даже во время движений крылом, и поэтому третичные перья у них либо отсутствуют, либо редуцированы до контурных перьев, подобных тем, которые располагаются на теле. Только специализированные на парящем полёте птицы, которые вытягивают крылья на всю их длину почти в прямую линию, имеют третичные перья, похожие по внешнему виду на вторичные маховые. Лишь у некоторых птиц известны третичные маховые перья, хотя они могли присутствовать у других эуорнитин, таких какIchthyornis, которые, вероятно, были чайкоподобными динамически парящими птицами. У более примитивных птиц, таких, как Archaeopteryx, третичные маховые перья отсутствовали, но имелись перья подобные контурным (а также лопаточные), частично заполнявшие промежуток между крылом и телом.
Промежутки между плечом и первичными, вторичными и третичными перьями крыла (которые вместе называются маховыми) были покрыты несколькими слоями более мелких перьев, известных как кроющие. У всех птиц, кроме самых примитивных, к малому пальцу крепятся контурные перья, формируя структуру, называемую крылышко, которая помогает в манёвренности и торможении.
Дополнительные перья, подобные кроющим, крепятся к плечу и называются лопаточными, они помогают заполнить промежуток между вторичными перьями и телом и, когда крыло сложено, частично покрывают маховые перья. При жизни, когда птица находится в состоянии покоя, лопаточные перья создают эффект плавного перехода большей части крыла в перья тела, вследствие этого обычно видны лишь кончики маховых перьев. У примитивных птиц этот эффект был гораздо менее выражен, так как их крылья были неспособны складываться и плотно прилегать к телу, а маховые перья были более заметными в состоянии покоя. У продвинутых эуорнитин крылья располагались высоко на теле, так, что передние конечности (плечевые кости) располагались параллельно друг к другу на верху спины. В результате сложенные крылья формируют своеобразный «плащ», окутывающий верхнюю часть тела птицы. У более примитивных птиц, многие из которых имели низко расположенные на теле крылья с плечами около груди, крылья складывались к бокам тела, оставляя контурные перья спины открытыми.
|
|
|
|
|
|
|
|
|
|
|
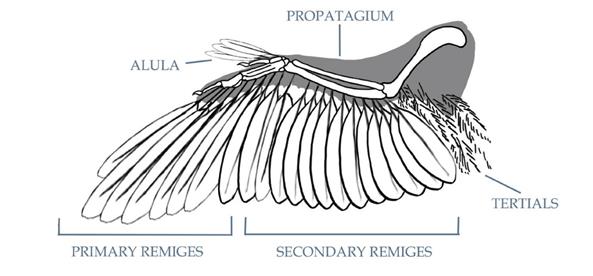
Вверху:Усреднённая анатомия крыла нептичьей авиаловой птицы.
У всех известных птиц, в том числе и у таких примитивных форм, как Microraptor, запястье соединяется непосредственно с плечом участком кожи и связок, известным как пропатагиум. Покрытая перьями и связанная с телом крыла, эта структура не позволяет локтю разгибаться в прямую линию и сглаживает передний край крыла, несмотря на то, что локоть постоянно удерживается под V-образном углом (что также поддерживает верхнюю часть крыла с большим или меньшим углом вдоль тела).
Общая форма крыла определяется длиной костей крыла и длиной маховых перьев. Большинство примитивных птиц имели широкие округлые крылья малого удлинения с относительно короткими первичными маховыми. Исключениями из этого правила являются Microraptor, Confuciousornis и Hongshanornis, у которых были очень длинные первичные маховые перья, придававшие их крыльям большое удлинение. Как правило, короткие и широкие крылья сочетаются с большей манёвренностью и часто встречаются у лесных птиц, которые летают короткими взрывными полётами и резко поворачивают между деревьев и среди кустарников. Длинные заостренные крылья часто встречаются у парящих птиц, таких, как чайки, или у тех, которые летают на высоких скоростях под открытым небом, как стрижи. Что характерно, большинство энанциорнисов, которые были почти исключительно древесными лесными птицами, имели короткие округлые крылья независимо от того, насколько продвинутым стал их анатомический летательный аппарат. Самые примитивные эуорнитины были наземными или водоплавающими птицами-универсалами, как современные куроподобные, сохранили сравнительно широкие округлые крылья, за исключением некоторых специализированных родов, таких, как Hongshanornis, которые летали в основном через открытые озёра и другие водоёмы. В этой книге для таких птиц, форма крыла которых неизвестна, например,Ichthyornis (морские птицы, реконструированные с длинными, заострёнными крыльями большого удлинения) приведена форма крыла с предварительной оценкой, основанной на их экологии и филогенетических отношениях.
Клювы и зубы
У современных птиц морда покрыта роговым клювом (известным в науке как рамфотека) и поэтому лишена перьев. Однако многие мезозойские птицы не имели клюва, вследствие чего степень оперения морды была у них очень разнообразна.Немногие окаменелости сохраняют тонкие и обычно короткие перья, присутствующие на морде, однако нам известны некоторые виды, например,Eoenantiornis buhleri и Sinornithosaurus millennii, у которых перья распространяются на три четверти длины к кончику челюстей или далее, что придаёт им курносый вид.У других видов, таких, как Microraptor zhaoianus и некоторых энанциорнисов («противоптицы»),имевших специализированные и удлинённые челюсти, перья не распространялись далеко вперёд от глаз.Вероятно, что, как и у современных птиц, у некоторых из них голова и/или шея были голыми.Лишённые перьев участки на голове у современных птиц обычно связаны с демонстрационным поведением, а в некоторых случаях улучшают теплоотдачу или облегчают уход за оперением.
Хорошо известно, что многие динозавры обладали клювом, но также известно, что многие из них вместе с клювом имели и зубы.Однако точное расположение клюва и зубов в челюсти обычно оценивается неверно.К примеру, почти любая реконструкция прижизненного обликаHesperornisимеетроговой клюв, покрывающий всю длину верхней и нижней челюсти.Некоторые из них явно демонстрируют зубы, прорезающиеся прямо из режущего края сплошного рогового клюва.Скорее всего, реконструкция сплошного клюва некорректна сама по себе, поскольку у всех не относящихся к классу Aves птиц, вероятно, имелась «составная рамфотека», т.е. клюв состоял из нескольких отдельных пластин, которые часто видны при жизни.
 У таких видов птиц, как Hesperornis regalis, нижнечелюстные зубы (зубной кости) занимают почти всю длину челюсти до её кончика, хотя самый кончик (и, вероятно, имевшаяся у этих птиц маленькая предзубная кость) был лишён зубов.На нижней стороне верхней челюсти (предчелюстной кости) имелись выемки, куда при смыкании челюстей помещались нижние зубы.Если бы присутствовал твёрдый клюв, то он должен бы был иметь углубления для размещения нижних зубов.Однако эти выемки присутствуют по краям челюсти. Края верхней челюсти слегка свисают на нижнюю челюсть, вследствие чего край клюва, если он имелся, не соприкасался с нижними зубами, что позволяло избежать стирания зубов при каждом смыкании челюстей.Расположение зубов у этого вида ограничено самой задней частью рта (верхнечелюстной кости). Об этом также можно судить по «зубным ячейкам» на нижней стороне черепа.
У таких видов птиц, как Hesperornis regalis, нижнечелюстные зубы (зубной кости) занимают почти всю длину челюсти до её кончика, хотя самый кончик (и, вероятно, имевшаяся у этих птиц маленькая предзубная кость) был лишён зубов.На нижней стороне верхней челюсти (предчелюстной кости) имелись выемки, куда при смыкании челюстей помещались нижние зубы.Если бы присутствовал твёрдый клюв, то он должен бы был иметь углубления для размещения нижних зубов.Однако эти выемки присутствуют по краям челюсти. Края верхней челюсти слегка свисают на нижнюю челюсть, вследствие чего край клюва, если он имелся, не соприкасался с нижними зубами, что позволяло избежать стирания зубов при каждом смыкании челюстей.Расположение зубов у этого вида ограничено самой задней частью рта (верхнечелюстной кости). Об этом также можно судить по «зубным ячейкам» на нижней стороне черепа.
 Согласно Heironymus & Witmer 2010 у Ichthyornis и Hesperornis предчелюстной ноготок и нижнечелюстной ноготок были наиболее сильно ороговевшими частями клюва. Эти «ноготки», часто формирующие небольшие крючки на кончиках клюва, находятся в зоне, где клюв должен быть наиболее прочным, как это наблюдается в клювах современных птиц.Те же авторы отмечают, что само наличие зубов на верхнечелюстной и зубной костях у этих видов, вероятно, означает, что у них полностью отсутствовали боковая роговая пластина надклювья и нижнечелюстная роговая пластина подклювья, которые обычно покрывают «губы» челюстей, и что наличие отвердевшей рамфотеки на краях челюстей может быть уникальной особенностью современных птиц.Однако, как было отмечено выше, кончик верхней челюсти (верхнечелюстная кость) у Hesperornis также не имел зубов, вследствие чего оставалось пространство для свисающего края, поэтому не исключено наличие некоего режущего края клюва.Должно быть, он был образован несколько более мягкой тканью, наподобие более гибкого клюва уток и гусей. Ещё одним доводом в пользу наличия клюва на предчелюстной кости является наличие «бороздки рамфотеки» на её верхней части спереди от ноздрей (носовых отверстий черепа), которая, вероятно, служила точкой фиксации рогового вещества на черепе.
Согласно Heironymus & Witmer 2010 у Ichthyornis и Hesperornis предчелюстной ноготок и нижнечелюстной ноготок были наиболее сильно ороговевшими частями клюва. Эти «ноготки», часто формирующие небольшие крючки на кончиках клюва, находятся в зоне, где клюв должен быть наиболее прочным, как это наблюдается в клювах современных птиц.Те же авторы отмечают, что само наличие зубов на верхнечелюстной и зубной костях у этих видов, вероятно, означает, что у них полностью отсутствовали боковая роговая пластина надклювья и нижнечелюстная роговая пластина подклювья, которые обычно покрывают «губы» челюстей, и что наличие отвердевшей рамфотеки на краях челюстей может быть уникальной особенностью современных птиц.Однако, как было отмечено выше, кончик верхней челюсти (верхнечелюстная кость) у Hesperornis также не имел зубов, вследствие чего оставалось пространство для свисающего края, поэтому не исключено наличие некоего режущего края клюва.Должно быть, он был образован несколько более мягкой тканью, наподобие более гибкого клюва уток и гусей. Ещё одним доводом в пользу наличия клюва на предчелюстной кости является наличие «бороздки рамфотеки» на её верхней части спереди от ноздрей (носовых отверстий черепа), которая, вероятно, служила точкой фиксации рогового вещества на черепе.
Так насколько же вытянутым был клюв?Heironymus & Witmer установили, что у птиц боковая роговая пластина надклювья почти всегда доходит до задней поверхности подноздревой перегородки. Этот отросток предчелюстной кости пролегает назад и отделяет ноздри от верхнечелюстной кости.Это означает, что клюв очень редко, если когда-либо вообще, налегал на саму верхнечелюстную кость.Верхнечелюстная кость у Hesperornis даже несколько компенсирует это ограничение, пролегая немного вперёд под подноздревой перегородкой, и удлиняет зубной ряд за пределы клюва.
Из приведённых выше свидетельств видно, что у Hesperornis беззубые, заострённые кончики клюва должны были состоять из прочного обычного кератина, тогда как остальная часть клюва была бы больше похожа на твёрдую кожу, ближе к черепу постепенно переходящую в нормальную кожу и перья.Ни при каких условиях зубы не могли занимать одно физическое пространство с рамфотекой, хотя они могли нахлёстываться друг на друга благодаря тому, что части зубного ряда размещались в челюсти.По-видимому, в рамфотеке никогда не было зубных ячеек, поэтому клюв и зубы по сути находились на разных частях челюстей и были отделены друг от друга.Hesperornis, пожалуй, является наиболее хорошо изученным примером птицы с клювом и зубами, но нет никаких оснований полагать, что те же общие принципы не будут верны для других птиц с клювом и без зубов.Короче говоря, ни одна мезозойская птица не имела «зубов в клюве», как это часто заявляется и изображается на рисунках.Скорее всего, клюв и зубы у них располагались в разных частях черепа, и, предположительно, выполняли различные функции в захвате пищи и её переработке.Зуб, растущий прямо из клюва, фактически приравнял бы роговое вещество к дёснам и в любом случае сделал бы клюв бесполезным.Зубы, растущие из клюва, были бы излишеством, которое не дало бы эволюционного преимущества.
С помощью филогенетического брекетинга возможно воссоздать приблизительную картину распространения клювов среди мезозойских птиц.Как представляется, клювов не было ни у одной из первых линий птиц, вероятно, потому, что и ранние авиалы (парящие птицы), и ранние дейнонихозавровые (птицы с серповидными когтями) были, в основном, хищниками, хотя некоторые из них, возможно, перешли к всеядности.Однако многие окаменелости дейнонихозавров, сохранившие перья, демонстрируют и маленький участок кончика морды, не покрытый перьями.Такой же лишённый перьев кончик морды наблюдается и у некоторых зубастых бесклювых энанциорнитин. Возможно, такое строение морды может быть признаком «рамфотеки» в самом широком смысле – очень слабо ороговевшей гибкой кожи, – как на задней части клюва у некоторых современных птиц, где рогоподобная кератиновая часть истончается и переходит в обычную кожу.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
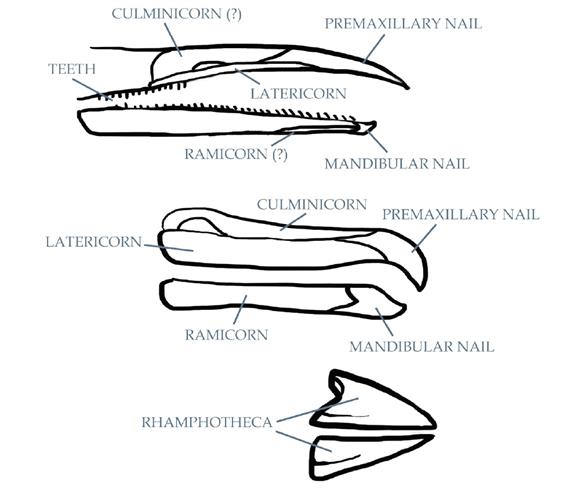
Вверху: Анатомические названия частей сложных клювов. Сверху вниз: клювы гесперорниса, альбатроса и вьюрка. Масштаб не соблюдён.
У нескольких независимых линий в основании клады Avialae общей тенденцией становится редукция зубов, вероятно, вследствие перехода к более всеядным рационам. Почти все известные базальные авиалы имели на верхней челюсти небольшое количество очень мелких зубов, а некоторые из них утратили зубы вообще.Эта тенденция, как оказывается, достигла своей кульминации у конфуциусорнитидов, которые не только не имели зубов, но и обладали остроконечными челюстями, а на некоторых очень редких образцах сохранился настоящий кератин клюва.На этих отпечатках видно, что у ранних обладавших клювом птиц, рамфотека была тонкой и хрупкой, и, вероятно, ороговевала гораздо меньше, чем у современных птиц.
У большинства энанциорнисов челюсти на всём протяжении несли зубы, а данные о наличии у них клюва отсутствуют.На основании этого может возникнуть желание объединить энанциорнисов и зубастых не имевших клюва дейнонихозавров в группу «Sauriurae», исключив из неё имевших клюв эуорнитин («истинных птиц»).Однако, учитывая, что у позвоночных много раз независимо возникал клюв, более вероятно, что каждый из представителей базальных авиалов с редуцированными зубами или вообще беззубый, возник независимо от других, или что у основания птицегрудых произошло обратное развитие, в результате которого у птиц снова появились челюсти, имевшие зубы на всём своём протяжении.В то время, как многие энанциорнисы сохраняют челюстной материал, только у одного вида наблюдается некая форма отсутствия зубов на передней части челюстей, что может свидетельствовать о наличии клюва: Gobipteryx minuta, как и конфуциусорнитиды, имел клюв и полностью утратил зубы.
Все известные мезозойские представители эуорнитин (вереерохвостых, к которым относятся и современные птицы), в отличие от типично бесклювых энанциорнисов, имели маленькие клювы, ограниченные кончиками челюстей, а также зубы в задней части челюстей. Несмотря на то, что хуншаньорнитиды изначально были описаны как имеющие клюв и абсолютно беззубые, O’Connor с коллегами позднее показали, что у них сохранялись ячейки зубов в верхней и, возможно, в нижней челюсти. Однако кончики челюстей были беззубыми и, вероятно, несли клюв.У некоторых (возможно, у большинства) мезозойских эуорнитинов перед зубной костью присутствовала дополнительная кость; эта предзубная кость никогда не имела зубов, и, вероятно, специально развилась для совмещения кончика челюстей и клюва.Предзубные кости известны у самых примитивных эуорнитин, таких, как Hongshanornis longicresta, до самых продвинутых нептичьих видов, например,Hesperornis regalis; однако, по-видимому, незадолго до появления современных птиц они были утрачены или же слились в одну твёрдую нижнечелюстную кость.Интересно, что единственная группа позвоночных, у которых также появились предзубные кости – это птицетазовые динозавры, имевшие сходное строение обладавших клювом и беззубых кончиков челюстей, располагавшихся перед полностью имеющими зубы челюстями.
Самые ранние эуорнитины, имеющие полноценный клюв и лишённые зубов, также относятся к самым примитивным: Archaeorhynchus не имели зубов, но обладали уплощённым клювом, похожим на клюв колпицы.Поскольку более продвинутые птицы сохраняли зубы на обеих челюстях, этот признак почти наверняка возник независимо, и не связан с отсутствием зубов у современных птиц.Сунлинорнитиды и поздние гесперорнисовые и Ichthyornis– все имели беззубую предчелюстную и предзубную кость, зубы сохранялись в верхнечелюстной и зубной костях. Исследования текстуры костей показали, что, вероятно, на кончиках челюстей у них имелся кератиновый клюв, а сзади от клюва – либо лёгкие челюсти с зубами, либо гибкая кожеподобная рамфотека.Поскольку обе главных линии современных птиц лишены зубов, вполне вероятно, что их общий предок также имел полноценный клюв, поэтому зубы, должно быть, были полностью утрачены в птичьей линии вскоре после появления ихтиорнисов. Интересно, что исследования структуры костей ихтиорнисов и гесперорнисов показывают, что они, вероятно, имели «сложную рамфотеку», и она могла быть наследственным признаком современных птиц (Heironymous & Witmer, 2010). Несмотря на то, что наиболее типичный птичий клюв состоит из одного покрывающего челюсти кератинового слоя, у видов со сложными клювами кератин на челюстях организован в отдельные пластины. Лучше всего это видно на примере некоторых современных морских птиц, например, у альбатросовых.
К сожалению, у таких интересных групп, как Hollanda, Gansus и Patagopteryxстроение нижних челюстей неизвестно. Однако мы можем использовать принцип экономии и филогенетический брекетинг, чтобы попытаться сделать обоснованный вывод. В большинстве исследований эти три группы относят к эуорнитинам («истинным птицам»), более примитивным по сравнению с гесперорнисами и Ichthyornis, имевшими верхние и нижние зубы и клюв на конце челюстей. Несмотря на то, что три промежуточные птицы снова могли независимо полностью или частично утратить зубы, при прочих равных условиях, вероятно, наиболее экономным вариантом для ихтиорнитид, гесперорнитид, хуншаньорнитид и чаояньорнитид было наличие клюва, ограниченного кончиками челюстей и зубов позади него.
Окрас перьев
Последняя работа Якова Винтера и его коллег по реконструкции прижизненного окраса доисторических птиц была одним из самых интересных палеонтологических исследований десятилетия. До этого исследования художники часто считали, что имеют полное право придумывать внешний вид динозавров. Однако, даже в отсутствие возможности напрямую увидеть цвета и оттенки у ископаемых видов, тем не менее существуют определённые биологические факторы, определяющие окрас птиц, которые в прошлом в значительной степени игнорировались художниками.
Имеются несколько процессов, придающих перьям цвет. На самом базовом уровне, они могут быть отнесены к структурному цвету, либо пигментации, хотя обе эти формы часто объединяются для создания прижизненного окраса птиц.
Структурные цвета получаются из реальной физической структуры кератина или меланина в пере. На микроскопическом уровне многие перья, демонстрирующие структурный цвет, имеют «пенистую» текстуру из мелких сфер или каналов, содержащих мелкие пузырьки воздуха. Особенности расположения этих пузырьков определяют характер рассеяния проходящего через них света. Развитие этих сложных структур недавно было рассмотрено Dufresne & al. (2009). С другой стороны, структура может быть получена путём наслоения или расположения в пространстве гранул меланина в пере (Stettenheim, 2000).
Структурные цвета оказывают двойное действие на прижизненный внешний вид – они создают цвета, отсутствующие среди различных пигментов, и расширяют или изменяют пигментные цвета. Например, среди амниотов (позвоночных, характеризующихся наличием зародышевых оболочек) нет ни одного известного способа синей пигментации. Синий окрас кожи, чешуи и перьев получаются при рассеивании света из-за структурной конфигурации. Подобным образом радужный перелив перьев, наблюдающийся у многих птиц, возникает из-за структуры пера. Птица с ярко-белыми или чёрными, как смоль перьями, вероятно, использует структурные цвета в дополнение к пигментам (или их отсутствию) для достижения подобного эффекта, без них эти цвета будут более равномерными, монотонными и менее яркими. Структурный окрас может действовать также и в качестве фильтра, изменяющего отражённый от пигментов свет и формирующего новые цвета. У большинства имеющих его птиц, зелёный цвет перьев получается благодаря наслаиванию жёлтых узелков пигментации на фоновую синюю структуру.
Несмотря на то, что в летописи окаменелостей трудно найти пигменты или химические следы, структурные цвета можно обнаружить в некоторых ископаемых перьях. Радужный перелив 45 ископаемых перьев описывался Vinther & al. (2008), и это иногда заметно даже невооружённым глазом. Структурный окрас перьев был распознан по отчётливой организации, в которой тонкий слой плотно упорядоченных гранул меланина покрывает более рыхлый конгломерат меланина. Её можно увидеть, даже если вышележащий разрозненный слой разрушен (Vinther & al. 2008). Подобное расположение, где структура является производным меланина, а также «пузырьков» в слое кератина, в первую очередь встречается в ослепительно радужном оперении колибри (Prum, 2006).
Механика структурного окраса перьев влияет на реконструкцию вымерших видов птиц. Синий, зелёный, чёрный как смоль, ярко-белый цвета не могут присутствовать у тех птиц, у которых отсутствует структурный окрас их перьев. Структурные цвета могли присутствовать или же не присутствовать в однонитевых перьях некоторых примитивных целурозавров и орнитисхий. Примечательно, что структурный окрас не наблюдается в однонитевых волосах современных млекопитающих. Однако основное отличие между волосом и простыми перьями заключается не в макроструктуре волокон, а в микроструктуре основополагающих молекул. Волос состоит из альфа-кератина – спиралеобразной молекулы наподобие ДНК. Бета-кератин, который входит в состав перьев, имеет слоистую и складчатую основополагающую молекулярную структуру, более подходящую для рассеивания света. С другой стороны, во всех ископаемых перьях, имеющих радужный оттенок, изученных Vinther & al. (2008), структурный окрас был ограничен бородочками, которых нет у многих примитивных пернатых динозавров. Кроме того, структурные цвета отсутствуют у современных пуховых перьев (пуха) и у пуховых придаточных перьев контурных перьев, в других частях имеющих структурный окрас. Поэтому вполне вероятно, что синие, зелёные, радужные, или яркие пуховые и однонитевые перья были крайне редки, если они вообще существовали у мезозойских птиц и более примитивных пернатых динозавров.
Подавляющее большинство оттенков окраса птиц полностью или частично определяется пигментацией или её отсутствием (Stettenheim, 2000). Существует несколько различных видов пигмента, наиболее часто встречающиеся из которых – меланины и каротиноиды.
Меланины легко определяются в ископаемых перьях, а их форма и концентрация может указать, какие цвета они создавали. Меланины отвечают за чёрный (но не глубокий, сплошной чёрный, который требует добавления структурного цвета), серый и большое разнообразие от коричневых до рыже-оранжевых и ржаво-красных цветов. Отсутствие меланина будет создавать белый окрас, что наблюдается у особей-альбиносов. Примечательно, что некоторые альбиносы современных птиц не совсем белые, а сохраняют некоторый тёмный окрас из-за структурных оттенков перьев, которые всегда сохраняются, даже несмотря на отсутствие меланина.
Каротиноиды, по большому счёту, являются тем, что придаёт птицам их характерные яркие цвета. У большинства животных каротиноиды не могут быть синтезированы непосредственно организмом (некоторые из них могут делать это, но для преобразования должны присутствовать другие типы каротиноидов). Каротиноиды появляются почти исключительно за счёт растительноядной диеты и, во вторую очередь, за счёт веществ, поглощающих множество каротиноидов в тканях их организмов (как растительноядные беспозвоночные и некоторые рыбы). У чаек, живущих вблизи лососевых ферм, перья приобретают розовый оттенок: это происходит потому, что выращенного на фермах лосося кормят искусственными источниками каротиноидов, чтобы сделать его мясо розовым, и это передаётся птицам. Самый необычный источник каротиноидов, на этот раз среди плотоядных видов, использует египетский гриф, который приобретает свою ярко-жёлтую кожу морды, поедая навоз копытных, который не даёт никакой существенной питательной ценности и, очевидно, потребляется грифами только из-за содержания каротиноидов (McGraw 2006). Действительно, в то время как хищники обычно не имеют яркой окраски, некоторые виды под действием селективного эволюционного давления могут вносить в свой рацион необычные добавки с целью стать более красочными (McGraw 2006).
 Каротиноиды часто используются как критерий соответствия при выборе партнёра. Поскольку каротиноиды должны поступать с пищей, птица с плохим питанием будет иметь более тусклый окрас, чем птица, которая очень успешна в поисках пищи. Содержащийся в зоопарке фламинго станет белым, если его диета не будет искусственно пополняться красными каротиноидами. Каротиноиды также могут повлиять на цвет глаз птицы, а также цвет клюва и цвет чешуек на её стопе: даже желтизна желтка куриного яйца (Zongker 2007) зависит от каротиноидов (некоторые птицы для окраски желтка используют флавин, который мы обсудим в этой главе позже).
Каротиноиды часто используются как критерий соответствия при выборе партнёра. Поскольку каротиноиды должны поступать с пищей, птица с плохим питанием будет иметь более тусклый окрас, чем птица, которая очень успешна в поисках пищи. Содержащийся в зоопарке фламинго станет белым, если его диета не будет искусственно пополняться красными каротиноидами. Каротиноиды также могут повлиять на цвет глаз птицы, а также цвет клюва и цвет чешуек на её стопе: даже желтизна желтка куриного яйца (Zongker 2007) зависит от каротиноидов (некоторые птицы для окраски желтка используют флавин, который мы обсудим в этой главе позже).
Примечательно, что даже современные птицы не имеют прецизионного контроля каротиноидной пигментации своих перьев. Каротиноиды почти всегда определяют окрас больших участков перьев, но не маленьких участков и не сложной детализации узора отдельных перьев(Hill 2010). Каротиноиды до сих пор не встречались в окаменелостях главным образом потому, что гранулы каротиноидов выглядят так же, как гранулы меланина (меланосомы), и, в отличие от меланина, каротиноиды нельзя отличить по форме гранул. Li & al. (2009) предложили провести специальные химические тесты для определения, является ли меланосома на самом деле каротиноидом, и какого цвета она была. Химический анализ цветовой структуры пера, проведённый Wogelius & al. (2011), приблизил нас на шаг ближе к возможности определения каротиноидного окраса ископаемых птиц.
 Даже если бы мы могли проверить наличие каротиноидов, остаётся неясным, могло ли большинство мезозойских птиц использовать их в качестве пигмента пера, как это делают современные птицы. Биологическая способность откладывать каротиноиды в перьях, очевидно, отсутствует у птиц подгрупп Palaeognathae (страусы, эму, киви, тинаму) и Galloanserae (включая уток, гусей, фазанов и т.д.). В то время как эти птицы могут использовать каротиноиды для окраски кожи ног, морды или клюва, им, как кажется, не хватает химических путей передачи каротиноидов в перья (Hill 2010).Если это не случай двух последовательных эволюционных инверсий, вполне вероятно, что красочные пигментированные каротиноидами перья являются уникальными для современной группы птиц Neoaves, и тогда почти все мезозойские птицы могли не иметь ярко-жёлтых, оранжевых, красных или зелёных перьев, но имели бы радужный и яркий контраст для создания эффективных визуальных демонстраций. Художники должны иметь в виду, что изображение оранжевых, жёлтых или зелёных перьев либо красных, оранжевых или жёлтых клювов или кожи, означает, что диета птицы содержит каротиноиды, и что даже этого могло быть недостаточно для обеспечения яркой окраски перьев птиц не относящихся к классу Aves.
Даже если бы мы могли проверить наличие каротиноидов, остаётся неясным, могло ли большинство мезозойских птиц использовать их в качестве пигмента пера, как это делают современные птицы. Биологическая способность откладывать каротиноиды в перьях, очевидно, отсутствует у птиц подгрупп Palaeognathae (страусы, эму, киви, тинаму) и Galloanserae (включая уток, гусей, фазанов и т.д.). В то время как эти птицы могут использовать каротиноиды для окраски кожи ног, морды или клюва, им, как кажется, не хватает химических путей передачи каротиноидов в перья (Hill 2010).Если это не случай двух последовательных эволюционных инверсий, вполне вероятно, что красочные пигментированные каротиноидами перья являются уникальными для современной группы птиц Neoaves, и тогда почти все мезозойские птицы могли не иметь ярко-жёлтых, оранжевых, красных или зелёных перьев, но имели бы радужный и яркий контраст для создания эффективных визуальных демонстраций. Художники должны иметь в виду, что изображение оранжевых, жёлтых или зелёных перьев либо красных, оранжевых или жёлтых клювов или кожи, означает, что диета птицы содержит каротиноиды, и что даже этого могло быть недостаточно для обеспечения яркой окраски перьев птиц не относящихся к классу Aves.
В то же время, более редкие, чем меланиновая и каротиноидная пигментация, пигменты порфирина обеспечивают другой способ получения цвета у птиц. Порфирины, – пожалуй, самые известные пигменты, придающие красный цвет крови и зелёный – листьям (в основу гема крови и хлорофилла входит порфирин), но они также могут окрашивать перья, добавляя коричневый, красный, а также зелёный цвет, хотя зелёный присутствует только в специализированной разновидности тураковердина, обнаруженной у тураковых. Интересно, что порфирины могут играть роль в терморегуляции. Вдобавок к теплоизоляционным свойствам яйца (см. ниже), они в основном встречаются в пуховых перьях ночных птиц, таких, как совы, и тех, которые активны при низких температурах. Другой причиной того, что порфирины встречаются в основном не в контурных перьях, является то, что это вещество делает перья менее устойчивыми, и поэтому было бы вредно использовать его в перьях, которые должны сопротивляться механическим нагрузкам и атмосферным явлениям. Это противоположно действию меланина, который увеличивает прочность и часто встречается на кончиках перьев крыла, где нагрузки являются наиболее высокими.
Порфирины часто ответственны за создание голубого окраса яиц странствующего дрозда и окраски большинства других яиц. И действительно, некоторые исследователи отмечают корреляцию между порфирином в яичной скорлупе и гнездовым поведением. Чисто белые яйца обнаружены только у птиц, которые гнездятся в таких укрытиях, как густая листва, и которые постоянно сидят на своих яйцах. Виды, которые оставляют свои яйца частично под воздействием атмосферных явлений, имеют окрашенную скорлупу, содержащую порфирин, отчасти ради маскировки, но, возможно, и в связи с предполагаемым терморегуляционным эффектом порфиринов.
Теоретически, порфирин можно обнаружить посредством химического анализа ископаемых птиц. Однако при реконструкции птиц без сохранившихся перьев, художники не должны злоупотреблять ещё не изученным меланиновым окрасом. Порфирины в перьях производят, главным образом, коричневый и тускло-красный – цвета, которые также могут быть произведены только меланином. Во всяком случае, порфирины дают художникам право добавлять дополнительные красноватые пятна чисто плотоядным видам, особенно таким, которые могли быть активны в ночное время или в условиях холодного климата.

Есть множество менее важных и более редких пигментов, которые могут окрашивать птичьи перья. Птерины отвечают за жёлтый, красный, белый и оранжевый цвет глаз некоторых птиц (у человека цвет глаз контролируется меланином, пониженное содержание меланина даёт эффект голубых глаз, с увеличением уровня меланина глаза у некоторых младенцев начинают темнеть). Флавиновые пигменты являются причиной жёлтого окраса многих яичных желтков. Пситтакофулвины встречаются только у некоторых попугаев, и создают жёлтые, оранжевые и красные цвета вместо каротиноидов, которые попугаи выделяют для поглощения, возможно, для пищевых целей. Есть (но в настоящее время не описаны) пигменты, известные только у пингвинов, которые придают флуоресценцию их жёлтым демонстрационным перьям.
Итак, окрас мезозойских птиц, должно быть, зависел преимущественно от меланина и радужных структурных цветов. Возможно, это делало окрас птичьей фауны более тусклым, чем мы наблюдаем сейчас, из-за сравнительно малого числа неоависов с их способностью окрашивать свои перья каротиноидами. Однако, как и современные утки, эти птицы всё равно могли бы создать сложные и ослепительно блестящие цвета и узоры, используя комбинацию естественных тонов меланинов (тускло-жёлтые, ржаво-красные, тёмно-серые и грязно-белые), а также многослойную переливчатость для создания ярких, подобных драгоценным камням, синих, зелёных, фиолетовых, глянцево-чёрных и белых оттенков. Тем не менее, несмотря на то, что они не могли бы иметь розовый окрас фламинго или ярко-зелёные и жёлтые цвета райских птиц, мезозойские птицы могли быть не менее красивыми.
Напротив: ИзображениеJeholornis prima.
Дата: 2019-07-24, просмотров: 536.