Дейнонихозавры, или «ящерицы со страшными когтями», в настоящее время представляют собой одну из самых примитивных известных линий птиц, обладавших хвостами с листовидными расширениями (вместе с родственной им линией Avialae). Окаменелости ранних дейнонихозавров относительно полны, а примитивные представители известны по ископаемым остаткам и отпечаткам перьев хорошей сохранности. Не удивительно, что примитивные представители каждой группы очень похожи и друг на друга и на примитивных авиалов, что свидетельствует о происхождении линии дейнонихозавров и линии, ведущей к современным птицам, от общего предка, очень похожего наArchaeopteryx lithogrpahica или Xiaotingia zhengi.
Примитивные дейнонихозавры, по всей видимости, были скользящими (т.е. способными к планирующему полёту),некоторые из них продвинулись к освоению активного машущего полёта.Microraptor zhaoianus имел хорошо развитые крылья и дополнительный комплект «задних крыльев», образованных из перьев с опахалом на нижней части ног и стопах, которые формировали бипланоподобную конфигурацию во время скольжения по воздуху или парашютирования с деревьев. У Archaeopteryx, Microraptor и Rahonavis крылья были достаточно большими и мощными, чтобы позволить неуклюжие взрывные полёты, хотя скользящий полёт, вероятно, был предпочтительным способом перемещения по воздуху.
За исключением ранних мелких планирующих видов, у дейнонихозавров наблюдается общая тенденция, прослеживаемая даже среди современных птиц: нелетающие наземные птицы становятся крупнее и более курсориальными (адаптируясь, таким образом, к наземному образу жизни). Эти более поздние и более крупные виды (представители группы Eudromaeosauria) включают в себя знаменитых «рапторов», которые на самом деле не были похожи на рептильных монстров, изображаемых в популярном кинематографе, но представляли собою очень крупных хищных наземных птиц. Некоторые из них обладали крыльями значительного размера (о чём свидетельствуют обнаруженные на костях крыльев Velociraptor mongoliensis бугорки, к которым крепились перья). «Рапторы» не очень похожи на своих кузенов карнозавров, скорее, они напоминают очень большого Archaeopteryx. Палеонтолог Марк Норелл в интервью по данному вопросу заявил:
«Чем больше мы узнаём об этих животных, тем больше мы понимаем, что в принципе не существует никакой разницы между птицами и их близкими динозавровыми предками, такими, как Velociraptor. Представители обеих линий имели вилочки, строили гнёзда, обладали полыми костями и были покрыты перьями. Если такие животные, как Velociraptor, в настоящее время были бы живы, на первый взгляд они производили бы впечатление странных птиц».
Даже характерно увеличенный «серповидный коготь» на втором пальце стопы большинства дейнонихозавров, вероятно, не служил оружием для удержания и убийства добычи. Его форма (и форма когтей передних конечностей) в большей степени напоминает когти лазающих животных. Способ крепления серповидного когтя и когтей крыла также идеально подходит для передачи нагрузки на остальные части стопы или крыла, в отличие от ожидаемой анатомии режущего оружия. Вполне вероятно, что эти когти сначала использовались небольшими планирующими видами для лазания по деревьям (наподобие того, как электромонтёры используют подъёмные клещи для брёвен), а позже были приспособлены более крупными наземными потомками для захвата добычи.
Подобная эволюционная тенденция развития от мелких планирующих к крупным наземным птицам у дейнонихозавров не является уникальной, но уникальным может оказаться их возвращение к хищническому образу жизни. Многие линии ранних манирапторов (птицы и их ближайшие родственники), очевидно, были всеядными, растительноядными или насекомоядными, и только среди эудромеозавров развилось гиперхищничество (рацион, включающий главным образом крупных позвоночных).
Тогда как дейнонихозавры, должно быть, возникли, по крайней мере, в средней юре, все известные примитивные представители этого семейства датируются поздней юрой или более поздней эпохой, и могли значительно отойти от формы тела предков, имевших, предположительно, маленькие размеры и большие крылья.
Первая известная мезозойская птица –Archaeopteryx lithographica была найдена в 1861 году и названа на основе единственного пера. Название было передано более полному скелету в 2011 г.Были распознаны несколько видов археоптеригид, найденные в известняках Золенгофена в Баварии. Однако большинство из них, вероятно, являются экземплярами A. lithographica в различных возрастных стадиях.Archaeopteryx традиционно считается более тесно связанным с современными птицами, чем с дейнонихозаврами, хотя вполне возможно, что на филогенетическом древе археоптеригиды находятся на дейнонихозавровой линии или базальнее разветвления авиалы/дейнонихозавры.
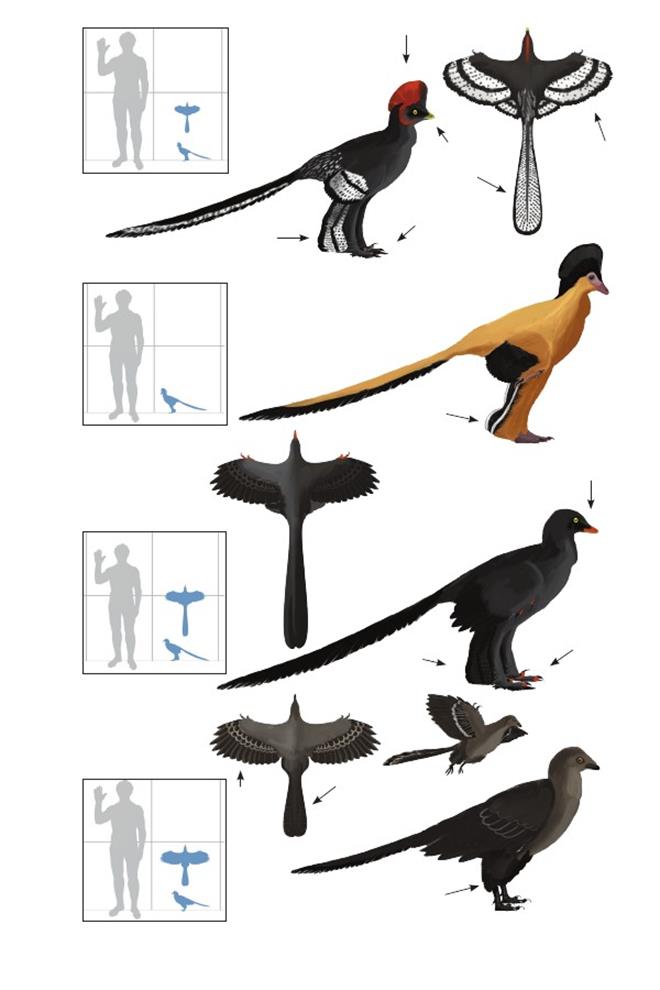
Почти птица Хаксли Anchiornis huxleyi
Время: 160 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания: формация Тяоцзишань.Размеры: РК 37 см (1,2 фута); ДТ 40 см (1,3 фута); ПД 45 см (1,5 фута).Характерные особенности: Голова треугольной формы с прямой заострённой мордой и выделяющимся красным/чёрным гребнем. Морда покрыта оперением почти до самого кончика. Контурные перья блекло-чёрного цвета с красными крапинками на морде. Крылья сравнительно короткие и округлые с узкими, симметричными маховыми перьями. Пальцы полностью покрыты перьями. Маховые и кроющие перья белые с ровными рядами чёрных крапинок на рядах первичных маховых и неровными чёрными пятнами на вторичных маховых. Ноги длинные с короткими задними крыльями. Перья задних крыльев усеяны чёрными пятнышками подобно передним крыльям, длинные рядом со стопой, ближе к туловищу сужаются. Большой, но слабоизогнутый серповидный коготь. Пальцы ног полностью покрыты перьями. Хвост очень длинный с крапчатыми рулевыми перьями, располагающимися до самого основания хвоста. Биология: Прижизненный окрас восстановлен по результатам исследований сохранившегося меланина, но могли существовать различия окраса между полами, возрастными стадиями или популяциями. Закруглённые крылья с неспециализированными маховыми перьями указывают на потерю способности к полёту, но были возможны парашютирование или ограниченный парящий полёт. Яркий окрас гребня и перьев крыла указывают на их роль в демонстрационном поведении.
Перо стопы из Даохугоу Pedopenna daohugouensis
Время: 155 млн. лет назад.Место находки: Внутренняя Монголия, Китай.Среда обитания: формация Даохугоу.Размеры: РК неизвестен; ДТ ~80 см (2,6 фута); ПД неизвестна.Характерные особенности: Известен только по стопам/нижней части ног. Ноги длинные. Задние крылья выражены, но редуцированы, с гораздо более длинными первичными маховыми перьями, чем вторичными. Первый палец очень тонок и не обращён назад. Биология: Как у Anchiornis, ножные перья сравнительно короткие и не имеют больших опахал, поэтому они не могли значительно влиять на аэродинамические характеристики.
Сяотин Чжена Xiaotingia zhengi
Время: 160 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания: формация Тяоцзишань.Размеры: РК ~60 см; ДТ 70 см; ПД неизвестна.Характерные особенности: Голова треугольной формы с удлинённой мордой. Крылья большие. Ноги большие и имеют задние крылья. Пальцы полностью оперены. Серповидный коготь на втором пальце стопы. Биология: Детали оперения неизвестны, вид реконструирован, как нечто среднее между A. huxleyi и A. lithographica.
Литографическое древнее крыло Archaeopteryx lithographica
Время: 150 млн. лет назад.Место находки: Бавария, Германия.Среда обитания: формация Золенгофен. Засушливый тропический остров в море Тетис. Низкие кустарники и пляжи, окружающие закрытые лагуны. Размеры: РК 77 см (2,5 фута); ДТ 55 см (1,8 фута); ПД 60 см (2 фута).Характерные особенности:Крылья широкие и округлые, с аэродинамическими перьями; по крайней мере, некоторые кроющие перья чёрного цвета. Пропорционально короткая часть пальцев с когтями выступает наружу. Короткие ноги покрыты длинными перьями, которые выходят за лодыжку. Хвост длинный с большим листовидным расширением из перьев, доходящих до основания хвоста. Биология: Примитивная анатомия крыла и относительно слабые стержни первичных маховых перьев указывают на неспособность летать. Изредка был возможен планирующий полёт, но, скорее всего, эти животные лишь бродили по пляжу. Большинство экземпляров мелкие (размером с голубя или меньше), а некоторые экземпляры были выделены в отдельные виды (такие как Archaeopteryx siemensi, Wellnhoferia grandis и Jurapteryx recurva), хотя все они, возможно, представляют собой различные возрастные стадии одного вида. В течение роста пропорции крыла и перьев оставались сравнительно постоянными.
Микрорапторы
Микрорапторы были самыми мелкими и среди самых примитивных орнитодесмидов, сохранившими наибольшее количество общих черт с археоптеригидами и ранними троодонтидами. Ранние представители этой группы по сравнению с другими примитивными птицами имели крупные размеры, однако поздние представители независимо вновь измельчали и обрели размеры, сравнимые с археоптеригидами. Никто из них не достигал умеренных размеров представителей других групп орнитодесмидов.
Микрорапторы, по всей видимости, были более хищными, чем большинство других ранних птиц, начав тенденцию, которая достигнет своего апогея у больших эудромеозавров.Большинство обладало длинными, узкими мордами с загнутыми и частично зазубренными зубами.Коготь на II пальце был больше, чем у археоптеригидов и многих троодонтидов тех же размеров, однако он остаётся по-прежнему относительно широким по сравнению с эудромеозаврами и мог служить в первую очередь инструментом для лазания. Существует свидетельство того, что даже самые мелкие виды, такие как Microraptor zhaoianus, иногда хватали добычу, которая приблизительно соответствовала их собственному размеру или же превышала его.
Предположение, что микрорапторы были ядовиты и обладали длинными торчащими клыкообразными зубами неверно и основывается на неправильной интерпретации зубов, которые вышли из своих ячеек во время фоссилизации.
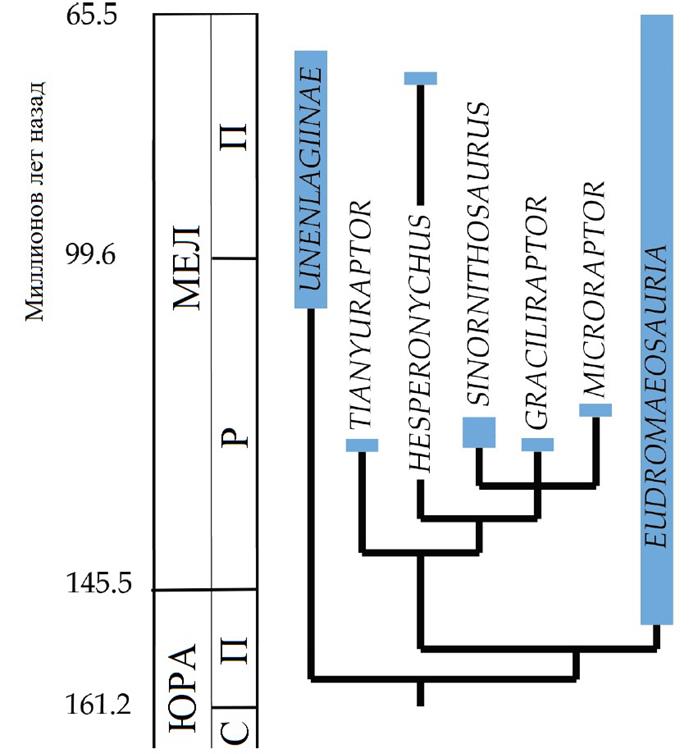
Вверху: Родственные связи микрорапторов во временной шкале. Филогения аппроксимирована на основе Senter et al. 2012.
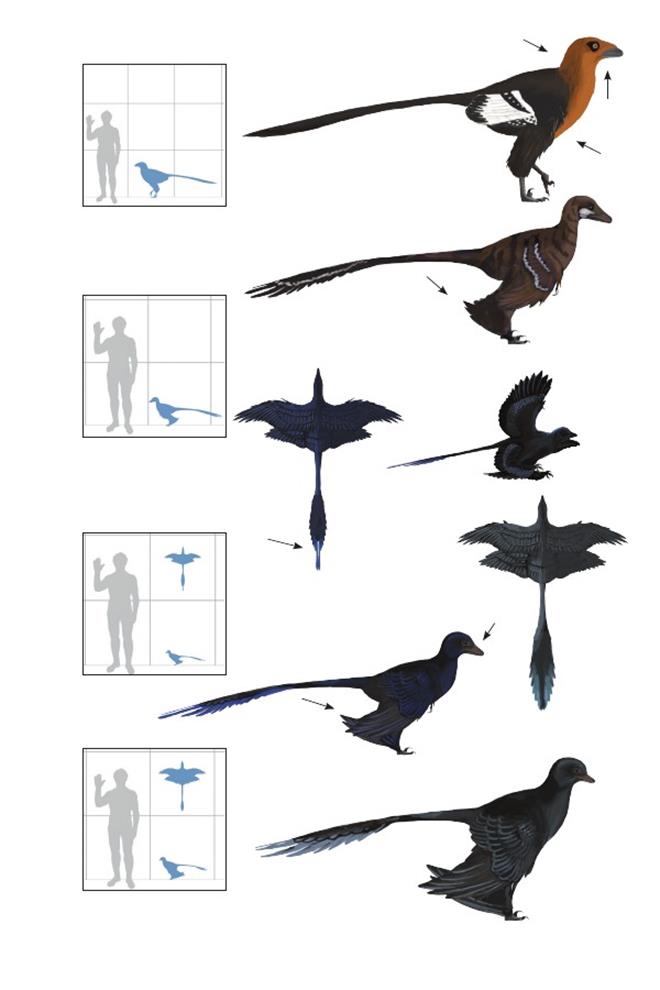
Хищник Тяньюй Острома Tianyuraptor ostromi
Время: 122 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания: Верхняя часть формации Исьянь (см. выше).Размеры: РК >60 см (2 фута); ДТ 1,7 м (5,5 фута); ПД неизвестна Характерные особенности: Морда широкая. Шея короткая и покрыта очень длинными перьями. Крылья маленькие, редуцированные. Ноги очень длинные. Хвост очень длинный и тонкий. Биология: Сильно редуцированная вилочка и длинные ноги указывают на полностью наземный образ жизни.
Мелкий хищник Ханьцина Microraptor hanqingi
Время: 122-120 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания: формация Цзюфотан, умеренно-субтропическая болотистая местность с преобладанием гинкго и хвойных деревьев, располагавшихся среди мелководных озёр и стоячих водоёмов. Размеры: РК 75 см (2,5 фута); ДТ 85 см (2,8 фута); ПД 90 см (3 фута).Характерные особенности: Очень похож на M. zhaoianus (внизу), но гораздо больше. Оперение плохо сохранилось. Маховые перья плюсны очень короткие по сравнению с экземплярами M. zhaoianus и C. pauli (приблизительно соответствуют длине костей плюсны). Возможная возрастная стадия M. zhaoianus; если это так, то это будет означать снижение способности к полёту даже по достижению половой зрелости.
Мелкий хищник Чжао Microraptor zhaoianus
Время: 122-120 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания: Исьянь (см. выше) и формация Цзюфотан, умеренно-субтропическая болотистая местность с преобладанием гинкго и хвойных деревьев, располагавшихся среди мелководных озёр и стоячих водоёмов. Размеры: РК 57 см (1,8 фута); ДТ 60 см (2 фута); ПД 68 см (2.2 фута).Характерные особенности:Голова большая. Морда узкая с зубами. Шея короткая. Первичные маховые перья длинные, аэродинамические, со слегка загнутыми назад стержнями. Вторичные маховые перья короткие, что делает крылья длинными и заострёнными с большим относительным удлинением. Ноги длинные с большими округлыми задними крыльями. Первичные и вторичные маховые перья задних крыльев почти равны по длине. Хвост очень длинный и тонкий с листовидным расширением из рулевых перьев в виде ромба, покрывающих его дистальную часть. У мужских особей последние два рулевых пера были удлинены. Оперение равномерно чёрного цвета с глянцевым радужным оттенком. Биология: Могли подниматься по стволам деревьев, цепляясь когтями крыльев, и прыгать с высоты. Листовидное расширение из рулевых перьев заканчивается двумя удлинёнными перьями, похожими на таковые у ранних авиалов, вероятно, они использовались в брачном демонстрационном поведении. Охотились на мелких энанциорнисов, но также хватали и более крупную добычу.
Скрытый летун Пола Cryptovolans pauli
Время: 75 млн. лет назад.Место находки: Альберта, Канада.Среда обитания: Формация Парк динозавров. Сезонно-засушливые низменные равнины с переплетённой системой рек и небольших лесов. Размеры: РК неизвестен; ДТ ~1м (3,3 фута); ПД неизвестна.Характерные особенности: Голова относительно небольшая, но морда неизвестна. Крылья хорошо развитые, длинные и заострённые. Грудина сросшаяся. Ноги длинные, содержат очень длинные задние крылья. Первичные и вторичные маховые перья задних крыльев почти равны по длине, кроющие перья очень длинные по сравнению с соответствующими маховыми. Задние крылья тянулись от основания пальцев стоп до бёдер, но, начиная с лодыжек, уменьшались в размерах. Хвост длинный с большим листовидным расширением из рулевых перьев в форме ромба, охватывавшем конечную половину его длины. Биология: Скорее всего, использовали планирующий полёт или слабо летали, задние крылья под передними крыльями образовывали форму биплана. Присутствие сросшейся грудины может свидетельствовать о возможности полёта. Задние крылья не обладают такими аэродинамическими свойствами, как передние крылья, но могли поочерёдно использоваться в качестве рулей или тормозов (Hall & al. 2012). Длинное хвостовое листовидное расширение, вероятно, использовалось для регулировки угла наклона (Habib & al. 2012). Описание гребня из перьев ошибочно, оно было основано на неправильной интерпретации раздавленной головы и оперения шеи. Microraptor gui – возможно, младший синоним, так как оба этих животных имели сросшуюся грудину.
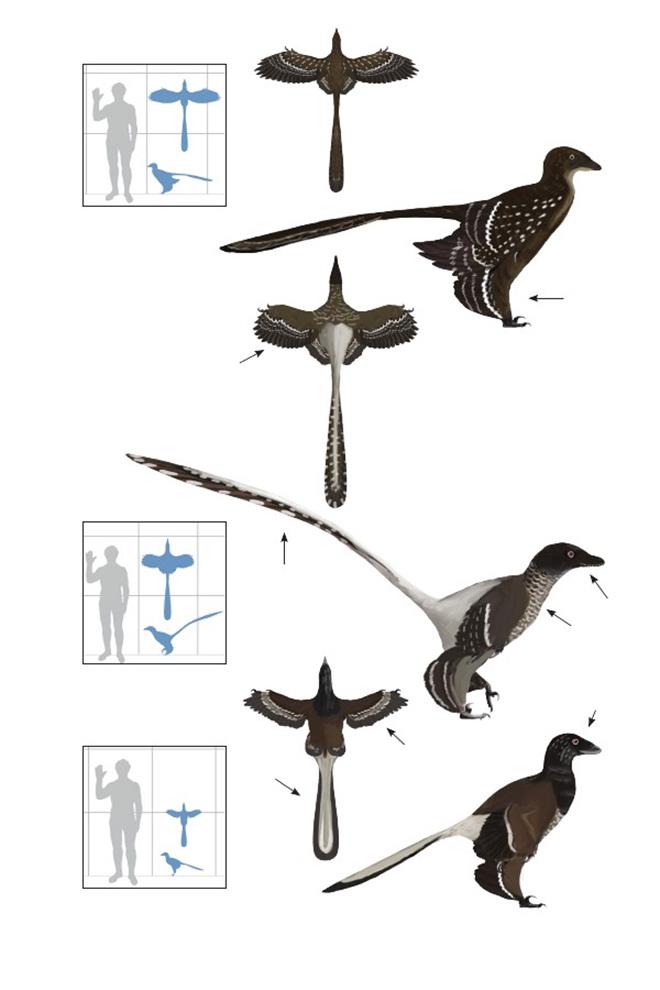
Грациозный хищник Луцзятунь Graciliraptor lujiatunensis
Время: 124,5 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания:Нижняя часть формации. Исьянь, умеренные хвойные и гинкговые леса, расположенные среди ряда озёр с притоками ручьёв и стоков с вулканических гор. Размеры: РК ~1,2 м (4 фута); ДТ ~1,1 м (3,6 фута); ПД неизвестна.Характерные особенности: Среднего размера. Крылья, вероятно, длинные и широкие. Ноги очень длинные, возможно, имели средние или большие задние крылья.Биология:Несмотря на крупные размеры, такие же, как и у его современника Sinornithosaurus, длинные ноги и тонкие пропорции сближали его с планирующими микрорапторами, и поэтому эти животные, вероятно, вели немного более древесный образ жизни, чем синорнитозавры.
Китайская птицеящерица тысячелетия Sinornithosaurus millennii
Время: 124,5 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания: Верхняя и нижняя часть формации Исьянь (см. выше).Размеры: РК ~1 м (3,3 фута); ДТ ~1,2 м (3,9 фута); ПД неизвестна.Размеры: (ювенильный экземпляр NGMC 91) РК 60 см (2 фута); ДТ 65 см (2,1 фута); ПД ~70 см (2,3 фута).Характерные особенности:У взрослых особей морда длинная и узкая, у молодняка – высокая и более треугольной формы. Перья доходят почти до кончика морды. Крылья редуцированы, симметричные маховые перья короче второго пальца. Ноги длинные, имеют длинные перья с опахалом, по крайней мере, на бедренной кости. Стопы чешуйчатые с большими когтями и серповидным когтем. Перья на плюсне неизвестны. Хвост очень длинный у взрослых особей и короткий у молодых, до самого основания покрыт перьями, имеющими опахала. Рулевые перья около кончика хвоста более крупные, образуют листовидное расширение в форме ромба. Очень пёстрый окрас, покрытие перьями тёмно-коричневого или чёрного и от светло-коричневого до желтовато-коричневого цвета.Биология: Похож на Graciliraptor, но с короткими ногами и меньшими крыльями. Маленькие крылья с симметричными опахалами перьев указывают на неспособность к полёту. Вероятно, сложная полосчатая окраска оперения служила целям демонстрационного поведения/маскировки.Изначально назван S. millenii, но согласно МКЗН, название должно быть исправлено на S. millennii.
Эудромеозавры
«Истинные дромеозавры», эти нелетающие орнитодесмиды, от средних до гигантских размеров, хорошо известны из позднемеловых отложений, но фрагментарные окаменелости и изолированные зубы свидетельствуют, что эта группа возникла, по крайней мере, в поздней юре.Их задние конечности были значительно редуцированы, что сильно ограничило скорость бега, но позволило развиться сильной мускулатуре, поддерживавшей серповидный коготь, использовавшийся для захвата и умерщвления крупной добычи.Сами серповидные когти были очень плоскими и гораздо больше подходили для удержания и разрывания, чем более широкие когти большинства других орнитодесмидов.Почти все они, как представляется, были гиперхищниками и специализировались на захвате более крупной добычи, чем они сами.На основании изучения следов и комплексов ископаемых остатков множества особей сделан вывод, что многие из них, вероятно, были стайными животными, мигрировавшими небольшими стаями.Как и у современных нелетающих птиц, относительно далёких от своих летавших или паривших предков, их перья, вероятно, вернулись к состоянию пухового пера с открытым опахалом, бородки которого в большинстве случаев не цепляются друг за друга, а крупные виды, обитавшие в жарких засушливых средах, могли совсем утратить перья на отдельных участках кожи для более эффективной терморегуляции.У современных птиц такое исчезновение перьев способствует теплоотдаче, но также может привести к перегреву.Зачастую лишённые перьев участки тела при необходимости могут частично или полностью закрываться перьями крыльев.
Несколько эволюционных линий эудромеозавров независимо друг от друга достигли крупных и гигантских размеров (аналогичная тенденция отмечена в эволюции гигантских уненлагиин рода Austroraptor и гигантских итемиринов, таких, как Itemirus).В обоих случаях эти гигантские виды обнаружены в отложениях раннего мелового периода; они могли конкурировать и, в конце концов, уступить дейнодонтидам ниши гигантских хищников, освободившиеся после исчезновения алозавроидов в северном полушарии.
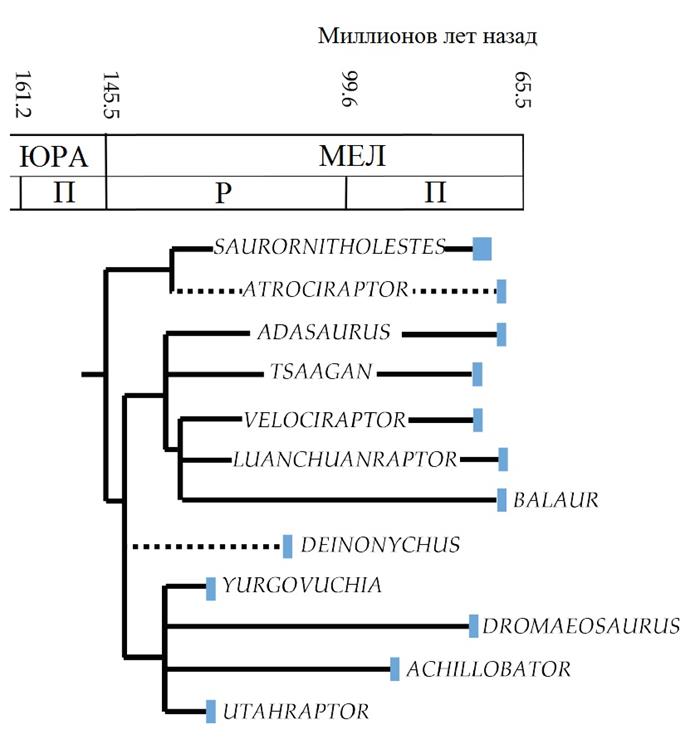
Вверху: Родственные связи эудромеозавров во временной шкале. Филогения аппроксимирована на основе Senter & al. 2012 и Turner & al. 2012
Хищник из Юты Острома и Мэя Utahraptor ostrommaysorum
Время: 126 млн. лет назад.Место находки: Юта, США. Среда обитания: стратон Йеллоу Кэт, формация Седар Маунтин. Открытые болотистые заиленные равнины. Размеры: РК неизвестен; ДТ ~6,5 м (21,3 фута); ПД неизвестна. Характерные особенности: Ноги короткие и сильные. Очень большие серповидные когти. Хвост относительно длинный.Биология: Один из самых крупных известных видов крылатых динозавров. Вероятно, хищник, специализировавшийся на крупной добыче, такой, как орнитоподы и молодые зауроподы.Крайне большие размеры могли быть приобретены из-за отсутствия в экосистеме крупных карнозавров после исчезновения в регионе аллозаврид, мегалозаврид и цератозаврид, что позволило эудромеозаврам занять ниши, обычно занимаемые другими тероподами. Появление в регионе кархародонтозавров (Acrocanthosaurus atokensis) совпадает с исчезновением U. ostrommaysorum.
Гигантский герой с ахиллесовым сухожилием Achillobator giganticus
Время: 90 млн. лет назад.Место находки: Дорноговь, Монголия. Среда обитания: формация Баин-Шире, вероятно, аналогичная более поздней формации Нэмэгэт – хорошо орошаемая, но засушливая полупустыня с преобладанием низкого кустарника, озёр и сухой лесистой местности. Размеры: РК >1,2 м (4 фута); ДТ ~5 м (16,5 фута); ПД неизвестна. Характерные особенности: Морда очень высокая и квадратная. Тело короткое и широкое с особенно длинной лобковой костью. Крылья маленькие, но с сильными когтями. Ноги очень короткие, что придавало приземистый вид. Хвост очень длинный. Биология: Мог пользоваться короткими, но сильными крыльями для удержания жертвы или в драках с соперниками. Ноги необычно короткие, голень и плюсна короче бедренной кости. Вероятно, являлся медленно передвигавшимся нападавшим из засады хищником.
Бегущий ящер из Альберты Dromaeosaurus albertensis
Время: 75 млн. лет назад.Место находки: Альберта, Канада. Среда обитания: формация Парк динозавров. Сезонно-засушливые низменные равнины с преобладанием системы разветвлённых рек и небольших лесов. Размеры: РК неизвестен; ДТ 1,7 м (5,6 фута); ПД неизвестна. Характерные особенности: Голова в профиль прямоугольная, с длинной квадратной мордой. Ноги по сравнению с современным ему S. explanatus относительно длинные. Хвост более гибкий, чем у большинства других эудромеозавров. Биология: Более длинные ноги, мощные челюсти и зубы указывают, что представители этого вида могли быть преследующими хищниками и охотились на более крупную добычу, чем другие современные им виды.
Дикий разбойник Маршалла Atrociraptor marshalli
Время: 70 млн. лет назад.Место находки: Альберта, Канада. Среда обитания: формация Каньона Подковы, сезонно-засушливые низменные равнины с разветвлённой системой рек и маленькими лесами. Размеры: РК неизвестен; ДТ ~1,7 м (5,6 фута); ПД неизвестна. Характерные особенности: Голова прямоугольная в профиль с короткой, округлой мордой. Биология: Вероятно, имел повадки, аналогичные повадкам Dromaeosaurus. Тяжёлый квадратный череп и массивные зубы указывают, что эти животные охотились на относительно крупную добычу.
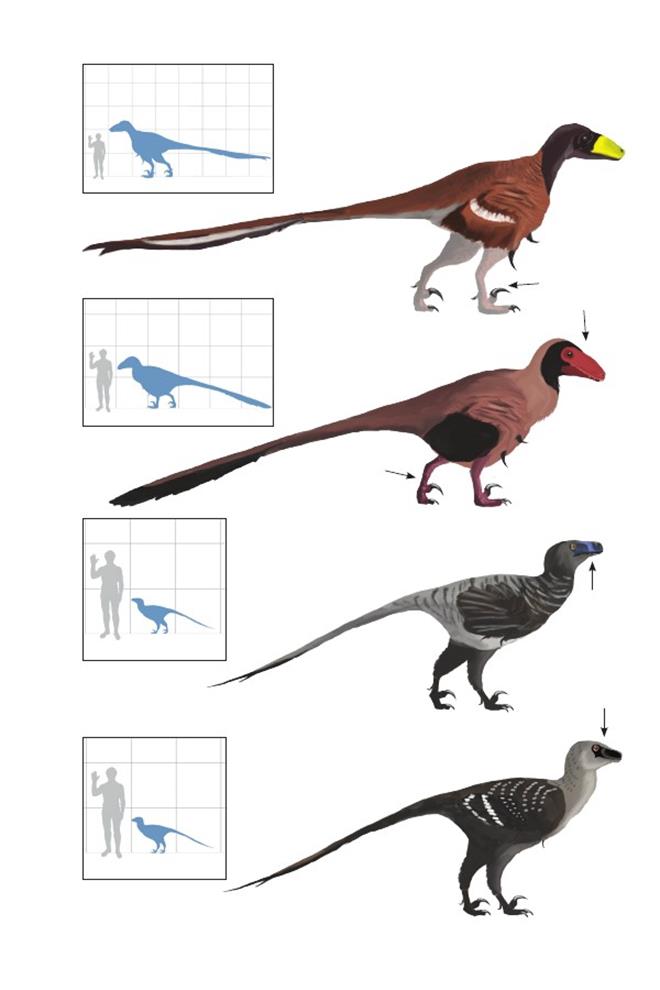
Койот Доелинга Yurgovuchia doellingi
Время: 126 млн. лет назад. Место находки: Юта, США.Среда обитания: нижний стратон Йеллоу Кэт, формация Седар Маунтин. Открытые заболоченные папоротниковые прерии, в которых были широко распространены игуанодоны, зауроподы, псевдозухи и полокантиды. Размеры: РК неизвестен; ДТ 2,4 м (7,9 фута); ПД неизвестна. Характерные особенности: Крупный эудромеозавр, известный по частичным скелетам. В положении покоя шея удерживалась относительно прямо (что следует издовольно-таки гладкой поверхности позвонков). Хвост более гибкий, чем у большинства других эудромеозавров.Биология:Один из нескольких эудромеозавров в этой экосистеме. Этот вид, очевидно, был наиболее крупным, его размеры приблизительно вдвое превышали размеры неназванных видов итемирин из той же формации. Возможно, является предком несколько более поздних гигантских дромеозаврин рода Utahraptor; достаточно толстая шея этих животных могла облегчать борьбу с относительно крупной добычей.
Балансирующий ужасный коготьDeinonychus antirrhopus
Время: 110 млн. лет назад. Место находки: Монтана, Оклахома и Вайоминг, США. Среда обитания: формации Кловерли и Антлерс. Тропические болота и поймы в дельтах рек, где преобладали хвойные, гинкго и древовидные папоротники. Покрытые в основном папоротниками и низким кустарником засушливые саванны с очень сухими сезонами.Размеры: РК >1,2 м (4 фута); ДТ 2,6 м (8,5 фута); ПД неизвестна. Характерные особенности: Высокая голова с высокой узкой мордой. Крылья сильно редуцированы, но сохраняют крупные когти. Неспособные плотно складываться крылья удерживались спереди или изгибались назад и укладывались вдоль боков. Ноги короткие и крепко сложенные, плюсна очень короткая. Крупные когти, большой серповидный коготь. Жёсткий хвост использовался, вероятно, для удержания равновесия. Биология: Из-за короткой плюсны, к которой крепились мышцы серповидного когтя, передвигался относительно медленно. Охотился на современных ему орнитоподовTenontosaurustilletti, в основномна молодняк. Возможно, охотился большими стаями. Молодые особи имели более длинные крылья и, возможно, вели частично древесный образ жизни. Серповидные когти молодых особей изогнуты ещё сильнее, чем у взрослых, и, возможно, использовались при лазании по деревьям.
Плоскийразбойникящер - птица Saurornitholestes explanatus
Время: 75 млн. лет назад. Место находки: Альберта, Канада. Среда обитания:нижние слои формации Парк динозавров/верхние слои формации Олдман. Сезонно-засушливые низменные равнины с разветвлённой системой рек и небольшими лесами. Размеры: РК >75 см (2,5 фута); ДТ 1,5 м (5 футов); ПД неизвестна.Характерные особенности: Крылья редуцированы, но молодые особи были способны захватывать мелкие объекты длинными противопоставленными пальцами. Морда короткая. Молодые особи отличались более массивной мордой и более длинными конечностями.Биология: Описание молодых особей основано на, вероятно, синонимичном виде Bambiraptor feinbergorum. Вид S. langstoni (Sues 1978), вероятно, является синонимом «Laelaps» explanatus (Cope 1876), последнее название имеет приоритет (Mortimer 2010).
Коренастый дракон Balaur bondoc
Время: 70 млн. лет назад. Место находки: Трансильвания, Румыния. Среда обитания: формация Себеш, часть острова Хацег моря Тетис. Климат тёплый, муссонный с горами, засушливые леса и озёра на возвышенностях и болотистые дельты рек в низинах. Размеры: РК >80 см (2,6 фута); ДТ ~1,5 м (5 футов); ПД неизвестна.Характерные особенности: Крылья большие. Третий палец утрачен, остальные пальцы крыла частично сросшиеся. Ноги короткие и мощные с серповидными когтями на первом и втором пальце.Биология: Первый палец стопы увеличен в размерах и поднят параллельно типично серповидному когтю. Необычно короткие и мощные задние конечности указывают то, что этот медленно передвигающийся хищник был приспособлен к охоте из засады и прижимал жертву к земле когтями.


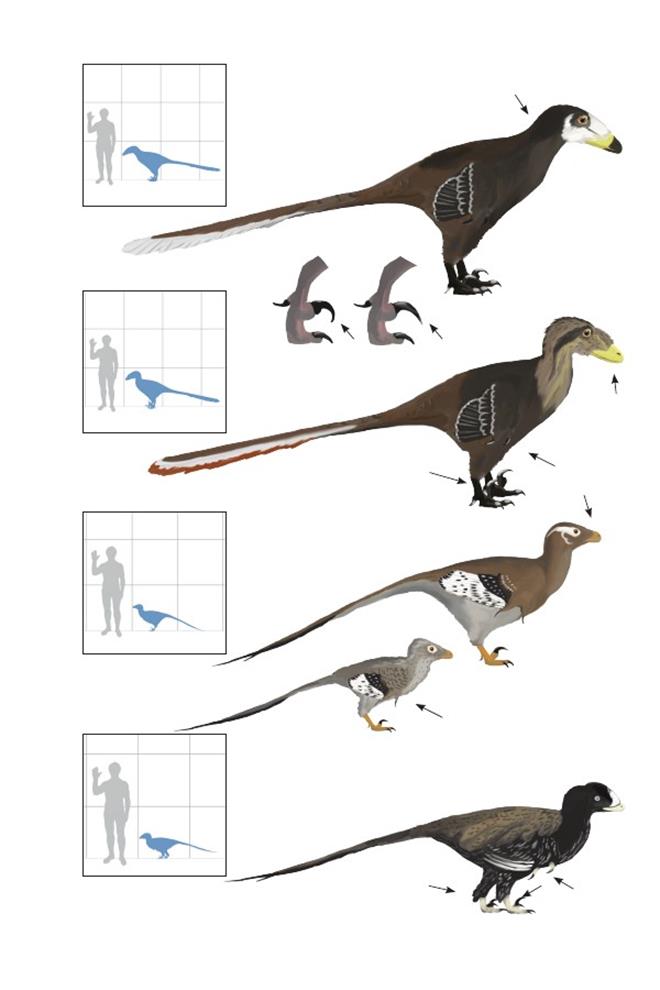
Монгольский быстрый хищник Velociraptor mongoliensis
Время: 75 млн. лет назад. Место находки: Умнеговь, Монголия и Внутренняя Монголия, Китай. Среда обитания: Байн-Дзак и Тугрикен Шире, формации Джадохта и Вулансухай. Пустынная возвышенность. Дюнные поля и сухие заросли кустарника. Размеры: РК >80 см (2,6 фута); ДТ 1,8 м (6 футов); ПД неизвестна.Характерные особенности: Морда длинная и немного вогнутая. Шея достаточно длинная. Крылья короткие с большими когтями и длинные вторичные перья с крепкими стержнями. Ноги короткие, с большим серповидным когтем. Тело широкое, особенно около таза. Хвост длинный и в некоторой степени гибкий.Биология: Охотился на превосходящих его размерами орнитисхий, в том числе Protoceratops andrewsi, также при случае питался падалью. Как и у Deinonychus, серповидный коготь молодых особей был изогнут сильнее, чем у взрослых. В отличие от других эуманирапторов, Velociraptor не обладал сильной мускулатурой, соединяющей плечо с верхней частью крыла, и поэтому не мог поднимать крылья выше горизонтальной плоскости (Parsons & Parsons 2009). На самом же деле, животные, вероятно, не могли поднимать плечевые кости даже до горизонтального положения. Подобное отсутствие подвижности крыльев, скорее всего, является вторичным признаком. Вид из Внутренней Монголии, названный V. osmolskae, отличается только пневматическими отверстиями черепа, строение которых у некоторых тероподов, как известно, изменчиво, поэтому, вероятно, он являетсясинонимом.
Вор из Луаньчуаня провинции Хэнань Luanchuanraptor henanensis
Время: не определено (поздний меловой период). Место находки: Хэнань, Китай.Среда обитания: формация Цюпа. Размеры: РК неизвестен; ДТ 2,7 м (9 футов); ПД неизвестна. Характерные особенности: Голова маленькая с обращёнными вперёд глазами. Шея умеренно длинная. Крылья маленькие, но массивные, сильные и мускулистые. Хвост длинный, тонкий и в некоторой степени гибкий.Биология: Формация Цюпа характеризуется такой же динозавровой фауной, что и Джадохта, в том числе гнездовьями овирапторид. По экологии, вероятно, аналогичен Tsaagan, хотя несколько примитивнее.
Белое чудовище Tsaagan mangas
Время: 75 млн. лет назад. Место находки: Умнеговь, Монголия и Внутренняя Монголия, Китай. Среда обитания: Укаа Толгод, формация Джадохта и формация Вулансухай. Дюнные поля и сухие заросли кустарника вблизи водоёмов. Места гнездовий большого количества различных пустынных птиц, в том числе ценагнатиформов, энанциорнисов и троодонтидов. Размеры: РК неизвестен; ДТ 1,2 м (4 футов); ПД неизвестна.Характерные особенности: Морда более высокая и более прямоугольная по сравнению с Velociraptor, зубы крупнее.Ноги длиннее, чем у других видов. Внутреннее строение черепа характеризуется наличием больших верхнечелюстных отверстий и отверстий на скуловых костях. Биология: Вероятно, предпочитал такую местную добычу, как овирапториды и их птенцы. Будучи современником близкородственного вида Velociraptor, должно быть, во избежание конкуренции специализировался на другой добыче или занимал другую нишу.
Монгольский ящер АдаAdasaurus mongoliensis
Время: 70 млн. лет назад. Место находки: Байянхонгор, Монголия. Среда обитания: формация Нэмэгэт. Хорошо орошаемые, но засушливые полупустыни с преобладанием низкорослого кустарника, озёр и сухой лесистой местности. Размеры: РК неизвестен; ДТ 2,7 м (9 футов); ПД неизвестна. Характерные особенности: Голова массивная, но строение морды не известно. Серповидный коготь очень маленький. Биология: Необычно маленькие серповидные когти указывают на уменьшение роли задних конечностей/когтей в захвате добычи.
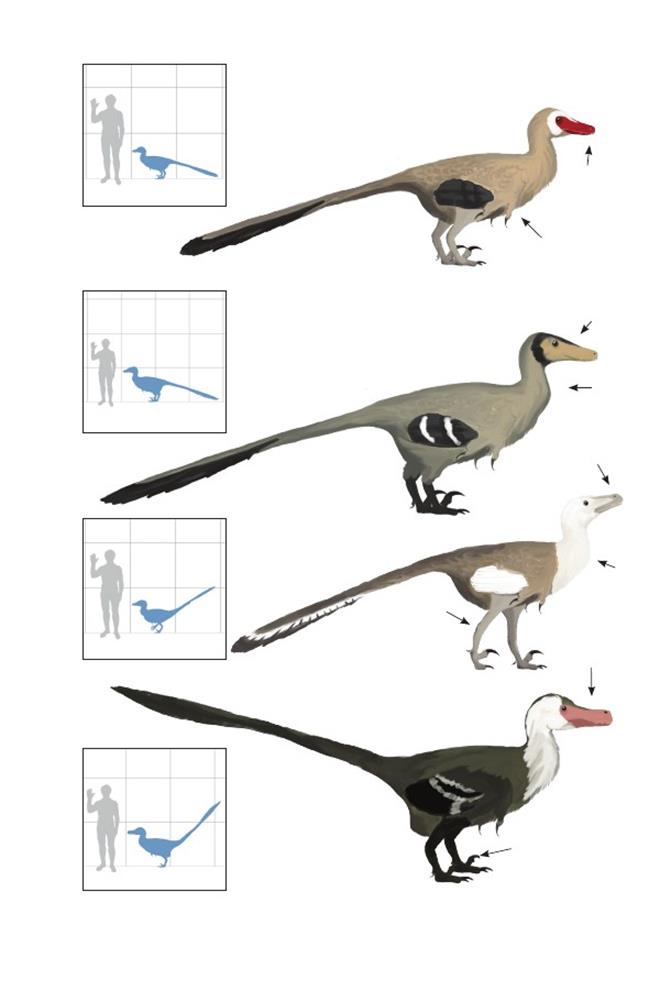
Уненлагиины
«Полуптицы»– узкоспециализированная группа бродивших по болотистому мелководью орнитодесмидов, наподобие цаплевых, приспособленных к засадной ловле рыбы. Они единственные птицы с листовидным расширением хвоста, обнаруженные южнее экватора и единственные длиннохвостые птицы южных континентов. Так как полёт окончательно сформировался у птицегрудых (энанциорнитины и эуорнитины), неизвестно, как представители этой группы смогли попасть в Южную Америку и на Мадагаскар.Возможный примитивный уненлагиин Rahonavis ostromi, несмотря на фрагментарность находок, демонстрирует хорошо развитые крылья и мог обладать достаточно активным полётом для перелёта на остров между северным и южным континентом, впоследствии дав начало распространению вторично нелетающих птиц-рыболовов. Как и у современных нелетающих птиц, перья нелетающих уненлагиин, вероятно, стали длинными и пуховыми (без сцепленного опахала).
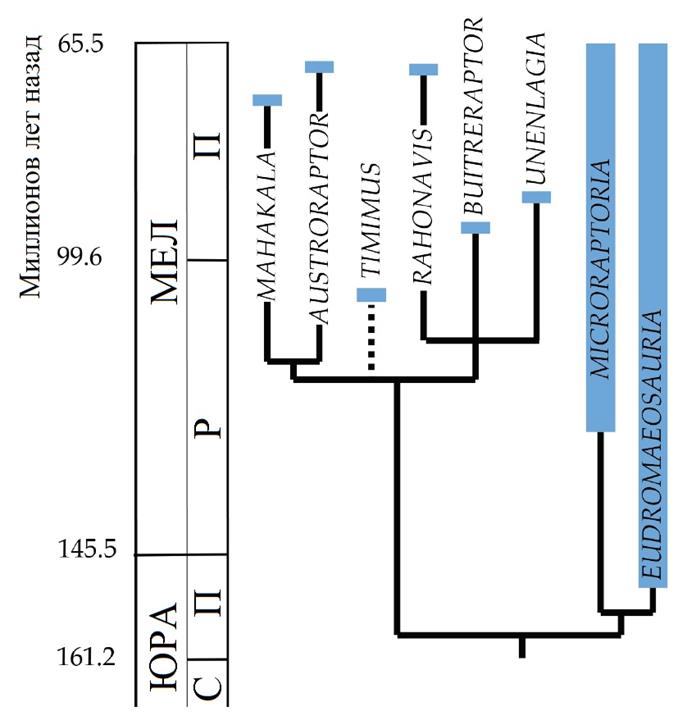
Вверху: Родственные связи уненлагиин во временной шкале. Филогения аппроксимирована на основе Senter 2007 и других источников.
Птица угроза-с-облаков Острома Rahonavis ostromi
Время: 70 млн. лет назад.Место находки: Махадзанга, Мадагаскар.Среда обитания: Формация Маеварано.Размеры: РК>80 см (2,6 фута); ДТ ~80 см (2,6 фута); ПД неизвестна.Характерные особенности: Очень маленький по сравнению с другими уненлагиинами и сравнительно примитивный, возможно, являющийся пережитком предковой формы. Крылья очень большие. Ноги длинные с необращённым первым пальцем и переразгибающимся серпообразным когтем второго пальца стопы.Листовидное расширение хвоста относительно короткое по сравнению с аналогичной структурой микрорапторов. Биология:Очень длинные, мощные крылья, имеющие маховые бугорки, позволяют предположить, что они были способны к некоторому ограниченному машущему полёту. Chiappe (2007) отметил, что, как и у других уненлагиин, крыло могло подниматься вертикальнее, чем у родственных групп, и позволяло делать им маховые движения. Примитивная активная стадия полёта представителей Unenlagiinae в целом может помочь объяснить их распространение в южном полушарии, которое в меловом периоде не сообщалось с северным материком. R. ostromi были одними из последних летающих длиннохвостых птиц.
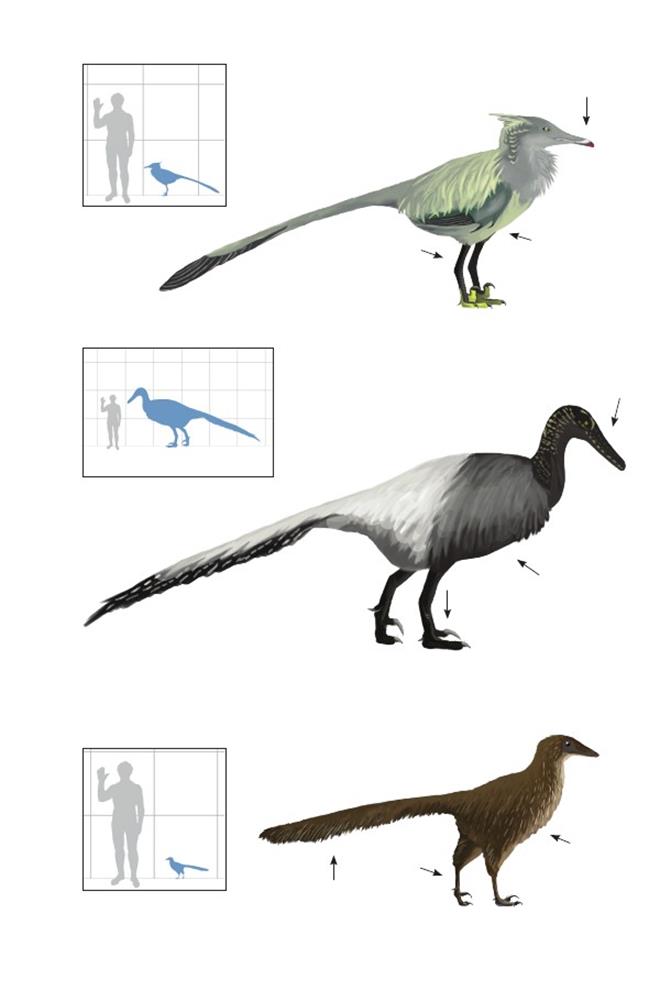
Германовский подражатель Тима Timimus hermani
Время: 106 млн. лет назад.Место находки: Виктория, Австралия.Среда обитания: Формация Эумералла, поймы и сплетения рек в умеренной и сезонно холодной антарктической окружающей среде. Размеры: РК неизвестен; ДТ ~3 м (9,8 фута); ПД неизвестна.Характерные особенности:Известен только по изолированным костям ног и позвонкам, принадлежавшим большому животному с длинными, тонкими ногами, приспособленными к бегу. Биология: Изначально рассматривался как орнитомимид, но, скорее всего, это эуманираптор, вероятно, уненлагиин.
Полуптица Комауе Unenlagia comahuensis
Время: 89 млн. лет назад.Место находки: Неукен, Аргентина.Среда обитания: Формация Портецуело.Размеры: РК неизвестен; ДТ ~3 м (9,8 фута); ПД неизвестна.Характерные особенности:Очень крупные нелетающие уненлагиины с маленькими, но очень подвижными крыльями. Внутренне строение характеризуется наличием большого выступа на дельтопекторальном гребне, поддерживающем сильную мускулатуру крыла. Биология: Экология, вероятно, схожа с экологией других представителей уненлагиин.
Полуптица Пайнемили Unenlagia paynemilli
Время: 89 млн. лет назад.Место находки: Неукен, Аргентина.Среда обитания: Формация Портецуело.Размеры: РК неизвестен; ДТ ~2,2 м (7,4 фута); ПД неизвестна.Характерные особенности: Похож на U. comahuensis, известен по фрагментарному материалу.Отличается некоторыми особенностями внутренней анатомии плечевого пояса, бедренных костей и, похоже, имел меньшие размеры. Биология: Экология, вероятно, схожа с экологией других представителей уненлагиин.
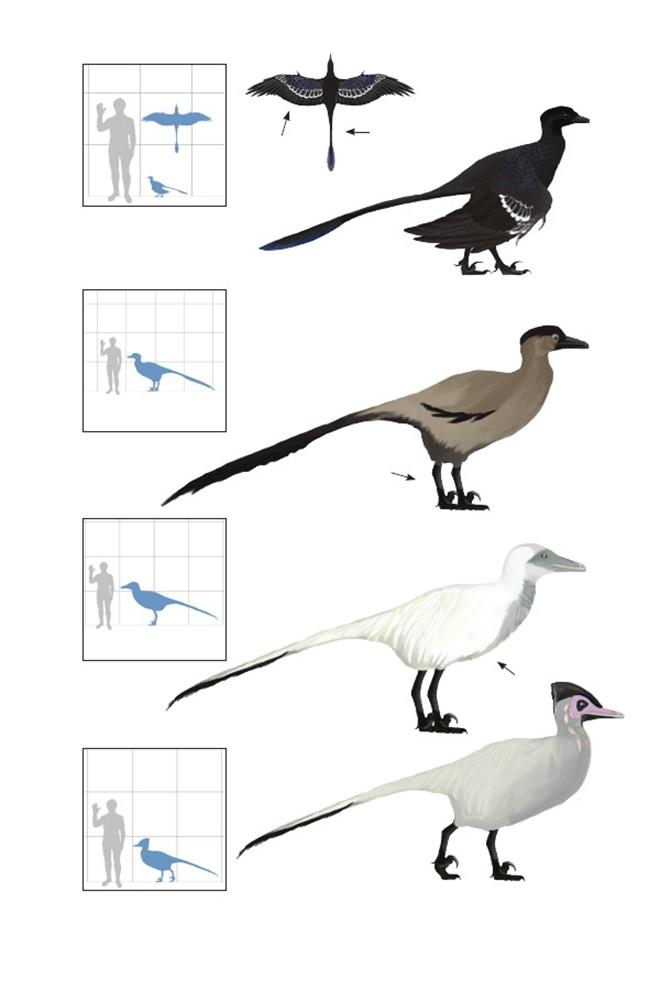
Хищник «насеста стервятника» Гонсалеса Buitreraptor gonzalezorum
Время: 94 млн. лет назад.Место находки: Неукен, Аргентина.Среда обитания: Формация Канделерос.Размеры: РК >70 см (2,3 фута); ДТ 1,3 м (4,4 фута); ПД неизвестна.Характерные особенности: Мелкие нелетающие уненлагиины со сравнительно большими крыльями. Морда чрезвычайно длинная и узкая. Когти крыльев короткие, а пальцы почти одинаковой длины.Кисть необычно короткая относительно очень длинных плечевых, лучевых и локтевых костей. Сравнительно длинные ноги. Биология: Очень длинная узкая морда предполагает рыбоядный рацион. Возможно, ходил и крался по мелководью для засады на рыб и мелких наземных позвоночных.
Южный хищник Кабазы Austroraptor cabazai
Время: 70 млн. лет назад. Место находки: Рио Негро, Аргентина.Среда обитания: Формация Аллен.Размеры: РК неизвестен; ДТ ~5 м (16,4 фута); ПД неизвестна.Характерные особенности: Гигантские нелетающие уненлагиины. Морда длинная и узкая с зазубренными зубами. Выступающий гребень над глазами. Крылья очень маленькие и, вероятно, в сложенном положении были плохо различимы среди оперения тела.Ноги длинные с плоским серповидным когтем на уплощённом втором пальце стопы.Биология:Вероятно, универсал, предпочитающий рыбу и озёрные берега, как современные цапли. Зазубренные зубы помогали хватать скользкую добычу. Серповидный коготь мог помогать разрывать более крупную добычу или туши. Вымирание спинозавридов, произошедшее на 10 миллионов лет раньше, могло позволить уненлагиинам, у которых была схожая экология, но гораздо меньшие габариты, приобрести более крупные размеры и занять пустые экологические ниши.
Махакала из УмнеговиMahakala omnogovae
Время: 75 млн. лет назад.Место находки: Умнеговь, Монголия.Среда обитания: Тугрикен Шире, джадохтская свита, сильно засушливая пустыня Гоби, кустарники и дюнные поля. Размеры: РК >20 см (7,8 дюйма); ДТ 65 см (2,1 фута); ПД неизвестна.Характерные особенности: Очень маленькие наземные дейнонихозавры. Крылья сильно редуцированы и, возможно, были не видны снаружи. Ноги тонкие. Хвост с виду короткий, но очень широкий и уплощённый.Внутреннее строение характеризуется уплощённой широкой локтевой костью и большим бедренным гребнем. Биология:Этот вид сохраняет некоторые признаки ранних дейнонихозавров, такие, как очень небольшие размеры, но другие особенности, например, чрезвычайно маленькие крылья, могут быть результатом обратной эволюции. Вероятно, промышляли мелкой добычей, обитавшей среди кустарников или зарывавшейся в песок. Эти птицы – одни из очень поздних наземных представителей этой линии (возможно, уненлагиин). Поскольку они собирали корм на земле, их оперение было реконструировано по аналогии с покровами современных мелких нелетающих птиц, таких, как киви.
Троодонтиды
Троодонтиды – «ранящие зубы»–сравнительно небольшая группа дейнонихозавров, в некоторых отношениях, впрочем, более похожие на современных птиц. Для них характерно более стройное телосложение с более длинными ногами, хорошо приспособленными для бега. Обычно они имели длинные узкие морды, несущие мелкие листообразные зубы, и хотя бы некоторые из них могли быть всеядными. Обычно они обладали меньшими крыльями и более короткими хвостами, чем другие крупные птицы с листовидным расширением из перьев на хвосте.
Продвинутые троодонтиды и троодонтины были похожи друг на друга по внешнему виду и размеру. Общая схема строения тела троодонтин оказалась очень успешной в позднем мелу, и в Азии, и в Северной Америке распространились очень сходные виды. Многие другие виды, наример, Pectinodon bakkeri, известны только по зубам, которые указывают на то, что троодонтиды просуществовали до самого конца мелового периода.
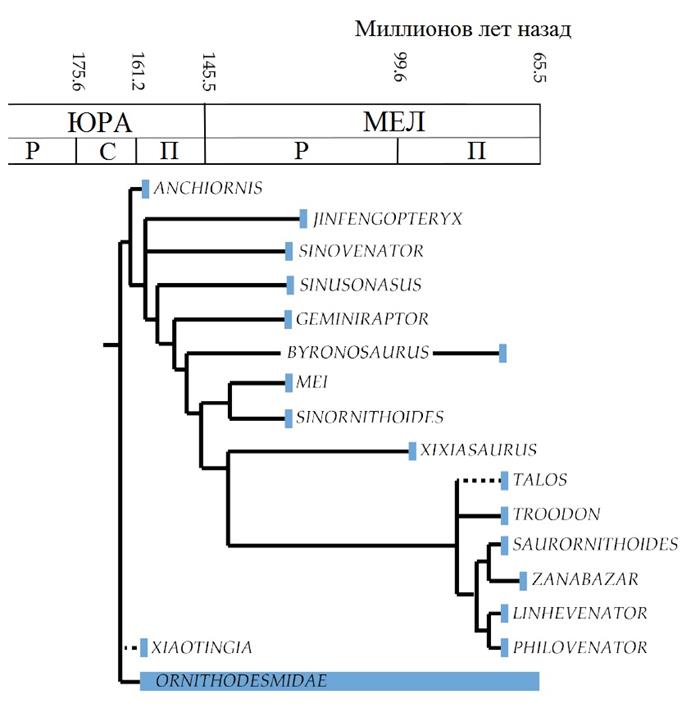
Вверху: Родственные связи троодонтидов во временной шкале. Филогения аппроксимирована на основе Senter et al. 2012.
Элегантное перо золотого феникса Jinfengopteryx elegans
Время: 122 млн. лет назад.Место находки: Хэбей, Китай.Среда обитания:стратон Цяотоу, формация Хауцзиин.Размеры: РК >35 см (1,1 фута); ДТ 55 см (1,8 фута); ПД ~65 см (2,1 фута).Характерные особенности:Голова треугольная, но округлая в профиль. Крылья маленькие с большими когтями. Туловище короткое. Длинные ноги лишены перьев. Хвост длинный, листовидное расширение из перьев доходит до его основания. Рулевые перья до ~10 см в длину на кончике хвоста и гораздо короче у его основания. Третий палец крыла редуцирован и сросшийся. Биология: Всеядный или растительноядный, кормился на земле семенами, орехами и, возможно, насекомыми и мелкими позвоночными.
Ящер из Хэнань Сися Xixiasaurus henanensis
Время: Не определено (поздний мел).Место находки: Хэнань, Китай.Среда обитания: формация Мацзяцунь Размеры: РК неизвестен; ДТ ~1,2 м (3,9 фута); ПД неизвестна.Характерные особенности:Морда длинная и округлая с U-образными челюстями (при взгляде снизу), слегка вогнутая по бокам. Биология: Крепко сложенный череп в форме коробки (а также большой общий размер тела) мог развиться как адаптация к поеданию крупной добычи.
Беспробудно спящий дракон Mei long
Время: 124,5 млн. лет назад.Место находки: Китай, Ляонин.Среда обитания:нижняя часть формации Исьянь, умеренные хвойные и гинкговые леса, расположенные среди ряда озёр, подпитываемых ручьями и притоком с расположенных неподалёку активных вулканических гор.Размеры: РК >30 см (11,8 дюйма); ДТ 45 см (1,5 фута); ПД неизвестна.Характерные особенности:Голова большая и округлая с короткой уплощённой мордой. Зубы многочисленные, мелкие, без зазубрин, расположены плотно, заполняют большую часть верхней челюсти, доходят до уровня глаз. Крылья относительно короткие. Ноги очень длинные с длинной плюсной и относительно короткой бедренной костью. Хвост длинный и гибкий. Внутреннее строение: вилочка U-образная, как у овирапторидов. Биология: Известен по экземплярам, найденным в спящих позах, вероятно, укрывавшихся от вулканического пепла, который похоронил их. В целом, похож на своего современника Sinovenator. Несмотря на ювенильный внешний вид имеет большую голову/глаза и длинные ноги, исследования гистологии кости показывают, что эти экземпляры были взрослыми (Gao & al. 2012).
Китайский охотник Чана Sinovenator changi
Время: 124,5 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания:Нижняя часть формации Исьянь (см. выше).Размеры: РК неизвестен; ДТ 1,1 м (3,6 фута); ПД неизвестна.Характерные особенности: Голова треугольной формы. Ноги длинные, крылья маленькие. Хвост длинный и гибкий, у некоторых экземпляров в спящей позе обёрнут вокруг тела.Биология:Вероятно, охотился на мелкую дичь вокруг озёрных систем Исьянь, но большинство окаменелостей было найдено в неозёрных зольных отложениях, что характеризует его, главным образом, как наземного обитателя внутренних областей суши. Похожий Sinusonasus magnodens является вероятным синонимом (Turner & al. 2012).
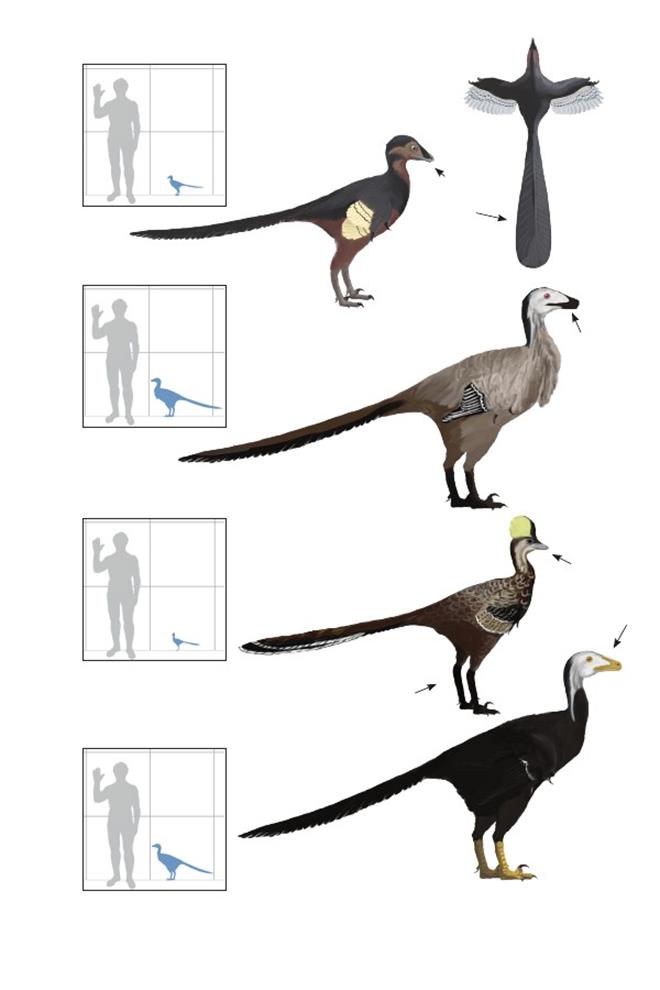
Захватчик-близнец Суареса Geminiraptor suarezorum
Время: 126 млн. лет назад.Место находки: Юта, США.Среда обитания: стратон Йеллоу Кэт, формация Сидар Маунтин. Открытые болотистые илистые равнины. Размеры: РК неизвестен; ДТ ~1,3 м (4,4 фута); ПД неизвестна.Характерные особенности: Троодонтиды средних размеров, известные по единственной кости морды (верхнечелюстной кости). Внутренняя часть высокой округлой морды характеризуется широкими воздушными полостями и выделяющимися, удлинёнными отверстиями.Ячейки зубов квадратные и разделены маленькими костяными стенками. Биология: Широкая морда и необычное расположение зубов может указывать на нестандартный для дейнонихозавров способ питания.
Китайская ящероптица Юнга Sinornithoides youngi
Время: 125 млн. лет назад.Место находки: Внутренняя Монголия, Китай.Среда обитания: формация Эджинхоро.Размеры: РК >50 см (1,6 фута); ДТ 1,3 м (4,4 фута); ПД неизвестна.Характерные особенности: Голова маленькая с несколько заострённой мордой. Шея сравнительно длинная. Длинное тело с очень маленькими крыльями и маленькими когтями крыльев. Ноги очень длинные с относительно маленькими когтями. Хвост сравнительно короткий.Биология:Чрезвычайно длинные ноги и очень маленькие крылья указывают на исключительно наземный образ жизни. Скорее всего, быстро бегали. Длинные и тонкие ноги, заострённая морда могут свидетельствовать о том, что они кормились в болотах.
Ящер Джаффе Байрона Byronosaurus jaffei
Время: 75 млн. лет назад.Место находки: Умнеговь, Монголия.Среда обитания: Укаа Толгод, джадохтская свита. Дюнные поля и засушливые лесистые районы с близлежащими водными протоками – места для гнездовья широкого разнообразия пустынных птиц. Размеры: РК неизвестен; ДТ ~1,4 м (4,6 фута); ПД неизвестна Характерные особенности: Морда длинная и узкая. Птенцы очень маленькие и по внешнему виду не похожи на родителей, имели заострённые головы треугольной формы. Гнездились неподалёку от ценагнатиформов (Citipati osmolskae) и энанциорнисов (Gobipteryx minuta) среди песка и дюнных полей. Биология:Возможно, охотились на таких мелких позвоночных, как ящерицы. Асимметричные ушные отверстия, как у сов, позволяли им точно определять местонахождение мелкой добычи, скрывающейся в кустах или зарывшейся в песок.
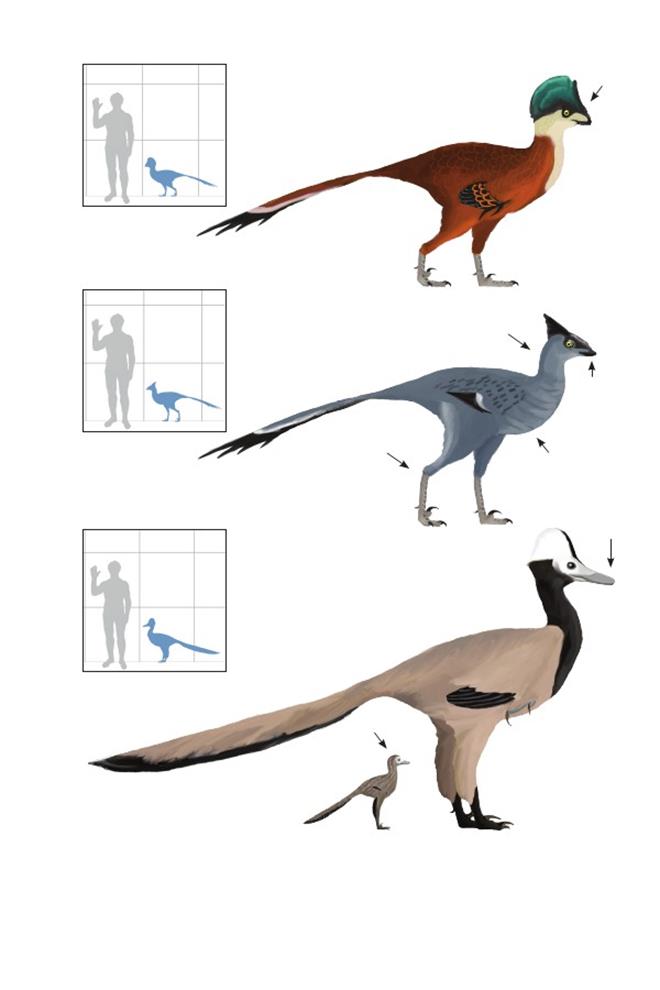
Монгольский птицеподобный ящер Saurornithoides mongoliensis
Время: 75 млн. лет назад.Место находки: Умнеговь, Монголия.Среда обитания: Баян Дзак, джадохтская свита. Засушливая пустыня, поросшая кустарником. Размеры: РК неизвестен; ДТ 1,5 м (4,9 фута); ПД неизвестна.Характерные особенности:Среднего размера по сравнению с видами той же эпохи. Морда длинная и узкая, но шире, чем у взрослого Byronosaurus. Зубы с большими грубыми зазубринами. Ноги длинные и тонкие. Биология: Длинные кости плюсны указывают, что они были быстрыми бегунами. Вероятно, питались мелкими позвоночными – ящерицами и выводками динозавров, добытыми в дюнах.
Молодой Занабазар Zanabazar junior
Время: 70 млн. лет назад.Место находки: Умнеговь, Монголия.Среда обитания: нэмэгэтинская свита (см. выше).Размеры: РК неизвестен; ДТ 2,5 м (8,2 фута); ПД неизвестна.Характерные особенности: Крупные троодонтиды. Морда длинная и узкая. Большое количество зубов (20 в верхней челюсти, 35 в нижней) с большими, крупными зазубринами. Ноги длинные и тонкие. Биология: Возможно, всеядные универсалы, предпочитавшие брать мелкую добычу, дополняя её растительным материалом, беспозвоночными и т.д.
Охотник Тань Линьхэ Linhevenator tani
Время: 75 млн. лет назад.Место находки: Внутренняя Монголия, Китай.Среда обитания: формация Уланьсухай.Размеры: РК неизвестен; ДТ ~1,7 м (5,6 фута); ПД неизвестна Характерные особенности: Морда сравнительно длинная, но череп широкий из-за расширенных скуловых костей. Крылья маленькие, но массивные и мощные. Серповидный коготь необычно большой для троодонтидов. Биология: Большой серповидный коготь развивавшийся, вероятно, параллельно у этого вида и у эудромеозавров, указывает на схожую стратегию захвата добычи у этих видов. Передние конечности короткие, как у других троодонтидов, но особенно мощные, возможно, использовались для копания, лазания или для захвата добычи.
Охотник Фила Карри Philovenator curriei
Время: 75 млн. лет назад.Место находки: Умнеговь, Монголия и Внутренняя Монголия.Среда обитания: формации Джадохта и Уланьсухай. Засушливая пустыня с кустарниками. Размеры: (ювенильный) РК неизвестен; ДТ ~75 см (2,5 фута); ПД неизвестна.Характерные особенности: Очень маленькие троодонтины. Голова треугольной формы с узкой заострённой мордой. Голова большая, ноги длинные и тонкие, и, что необычно, продольный размер ног больше поперечного. Серповидный коготь необычно маленький. Биология:Несмотря на то, что известен только по почти взрослым (до двух лет) экземплярам, часть из которых первоначально рассматривались как ювенильные Saurornithoides mongoliensis, они выделяются по нескольким анатомическим признакам, включая длинный листовидный отросток на большеберцовой кости, предположительно, для крепления мощных ножных мышц. Они, как представляется, более тесно связаны с Linhevenator. Другие экземпляры из того же временного промежутка и географической области, известные как «Zos Canyon Troodontid», вероятно, также относятся к этому виду (Mortimer 2010). Маленький серповидный коготь мог указывать на меньшую зависимость захвата добычи от когтей стопы.
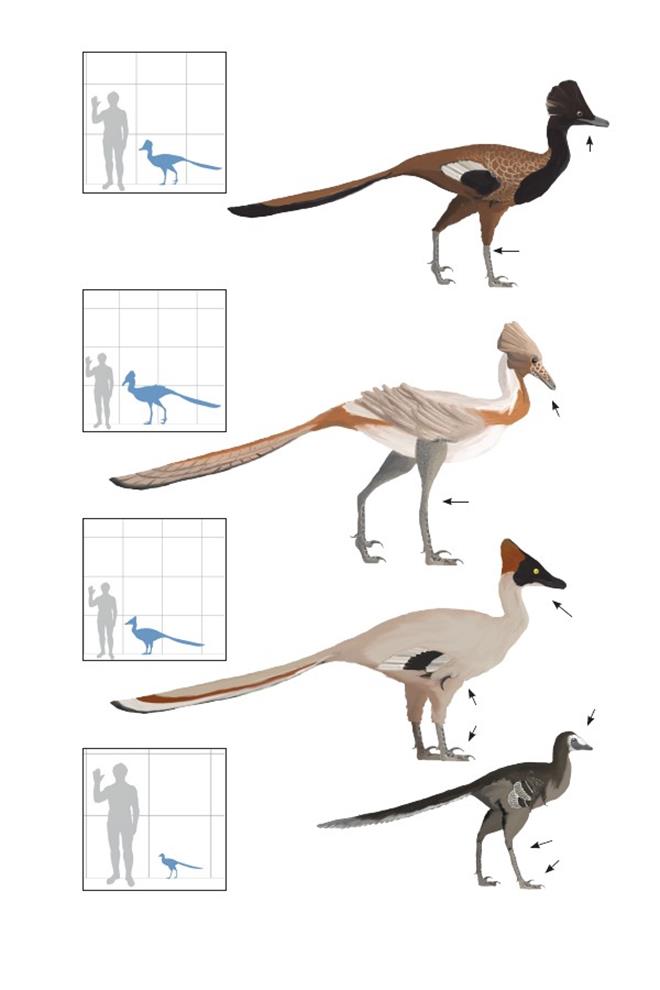
Талос СемпсонаTalos sampsoni
Время: 76 млн. лет назад.Место находки: Юта, США.Среда обитания: формация Каипаровиц. Пойма с преобладанием больших сезонно затопляемых рек. Занимала широкие илистые и песчаные берега рек. Размеры: РК неизвестен; ДТ ~1,6 м (5,2 фута); ПД неизвестна.Характерные особенности: Троодонтины средних и крупных размеров. Крылья очень короткие с тонкими предплечьями. Ноги тонкие, но короче, чем у азиатских видов. Серповидный коготь относительно маленький. Биология: Как и другие троодонтиды, эти птицы могли использовать свои большие изогнутые когти для захвата добычи, задействуя при этом, вероятно, одну стопу. Повреждение второго пальца стопы у первого известного ископаемого экземпляра подтверждает предположение, что палец стопы обычно использовался для прижатия к земле мелких, сопротивляющихся животных.
Красивый ранящий зуб Troodon formosus
Время: 75 млн. лет назад.Место находки: Монтана, США и Альберта, Канада.Среда обитания: формации Джудит Ривер, Парк Динозавров и Каньон Подковы. Сезонно засушливые и низменные равнины с преобладанием сплетений речных систем и небольших лесов. Размеры: РК неизвестен; ДТ ~2,3 м (7,5 фута); ПД неизвестна.Характерные особенности:Крупные троодонтины. Голова длинная, с длинной, но широкой мордой. Крылья очень маленькие. Ноги умеренно длинные, но более толстые, чем у азиатских видов той же эпохи. Серповидный коготь большой и уплощённый, похож на таковой у эудромеозавров, вероятно, использовался в хищническом поведении. Биология: Хищные, но, возможно, разбавляли рацион фруктами или семенами. Мелкие зубы, классифицированные как принадлежавшие видуParonychodon lacustris, возможно, принадлежали молодым T. formosus.Несмотря на то, что окаменелости, приписываемые T. formosus были найдены в огромном кругу формаций (включая очень крупные экземпляры с Аляски) и охватывают много миллионов лет (до 66 миллионов лет назад), большинство из них основаны на крайне фрагментарных остатках, и почти наверняка относятся к различным видам.
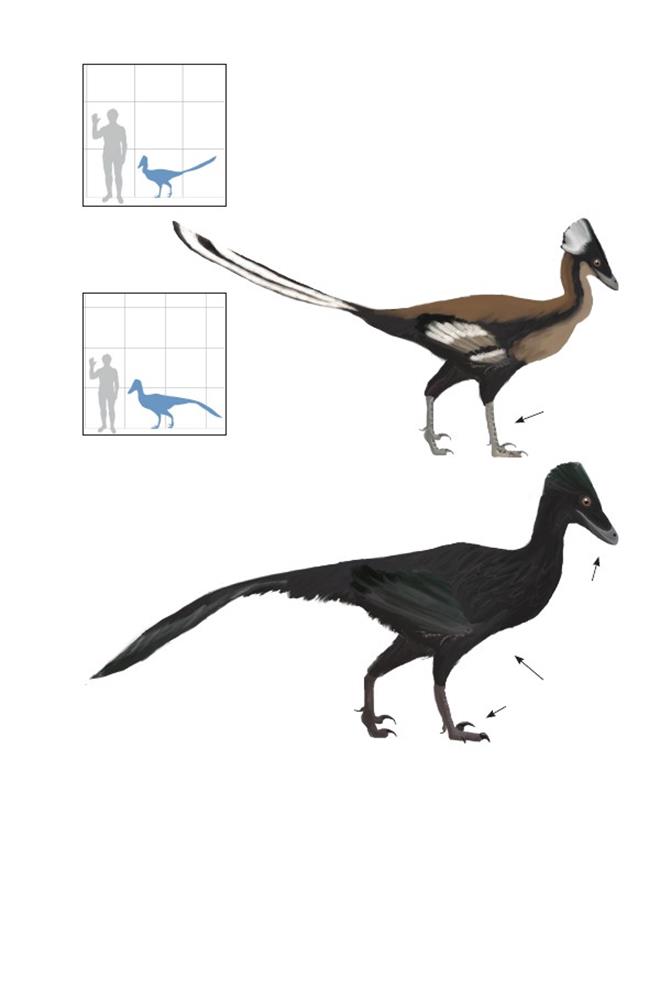
Базальные авиалы
Эта эволюционная линия, ведущая к современным птицам, вероятно, отошла от линии дейнонихозавров в течение средней или ранней юры. Все птицы более близкие к представителям класса Aves, чем к дейнонихозаврам, называются авиалами («крылатые птицы»), хотя это название используется также для группы, основанной на физическом признаке (наличие крыльев, используемых для полета), и поэтому может быть заменено на ФилоКод.
Успешная группа авиалов приобретала всё больше и больше признаков современных птиц, такие, как сросшиеся кости крыла, удлинённое относительно задних конечностей предплечье и укороченный хвост.Известны лишь немногие из этих более продвинутых длиннохвостых птиц, включая китайские формы Jeholornis и Yandangornis.Самая известная продвинутая ископаемая птица, которую уверенно можно считать «длиннохвостой», называется «Zhongornis haoae», имела сильно укороченный хвост, хотя и не столь короткий, как у Confuciousornis, у неё отсутствовал пигостиль или любое другое срастание хвостовых позвонков. Вероятно, экземпляр «Zhongornis» – всего лишь ювенильный конфуциусорнитид; этот факт могло бы стать любопытным свидетельством эволюции короткого хвоста: наличие длинного хвоста у цыплят короткохвостых птиц, как у головастиков, утрачивающих хвост по мере превращения в лягушку.
Авебревикауды – «короткохвостые птицы», являются первыми птицами, число хвостовых позвонков у которых сократилось до десяти или менее. Несколько последних позвонков обычно срастались вместе в единую прочную структуру, напоминающую пигостиль настоящих птиц (эуорнитин). Однако нет никаких свидетельств того, что эти хвосты обладали подвижным веером из перьев, наоборот, самые известные примитивные авебревикауды, по всей видимости, имели лишь несколько пар лентообразных рулевых перьев, если таковые вообще имелись.
Конфуциусорнитиды и другие примитивные короткохвостые птицы имели преимущественно длинные крылья, хотя у них не было хорошо развитой грудной мускулатуры, и они не могли поднимать крылья очень высоко над спинами, что препятствовало сильным взмахам и не позволяло им взлетать с земли. Небольшие обращённые внутрь и вперёд первые пальцы стоп, а также большие и сильные когти крыльев указывают, что вместо этого, с целью подняться на высоту, необходимую для планирующего или активного полёта, они карабкались на деревья. Многие виды обладали беззубыми клювами, которые развились независимо от таковых у современных птиц.
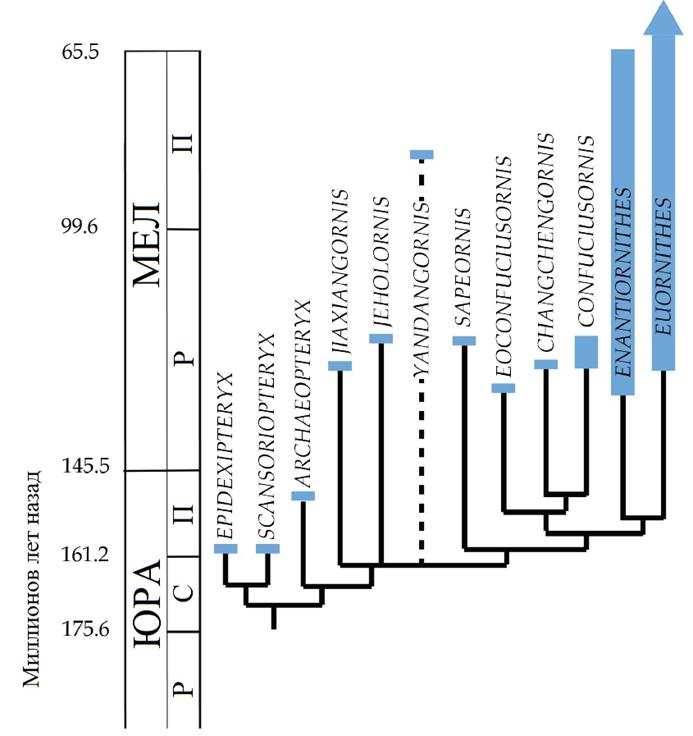
Вверху: Родственные связи авиалов во временной шкале. Филогения аппроксимирована на основеO’Connor & Zhou 2012.
Крыло для карабканья Гайлмана Scansoriopteryx heilmanni
Время: 155 млн. лет назад.Место находки: Внутренняя Монголия и Ляонин, Китай.Среда обитания: формация Даохугоу.Размеры: (ювенильный) РК>10 см (3,9 дюйма); ДТ 21 см (8,3 дюйма); ПД 22 см (8,6 дюйма).Характерные особенности:Голова округлая с короткой округлой мордой. Выступающие вперёд зубы. Крылья большие, с чрезвычайно длинным третьим пальцем. Ноги короткие, с четырьмя направленными вперёд пальцами. Хвост длинный, с листообразными перьями на кончике. Биология:Известен только по экземплярам выводковых птенцов. Обладал необычным набором как примитивных, так и продвинутых черт, которые сначала привели некоторых учёных к выводу, что эти животные не только являлись предками всех птиц, но были более примитивными, чем большинство динозавров. Этот вывод делался главным образом из-за отсутствия характерной для всех динозавров прободенной вертлужной впадины, что, однако, может быть объяснено находками ювенильных экземпляров. Малый палец был у них длинней, чем большой, что отличало их практически от всех динозавров. Удлинение третьего пальца с когтем, выступавшим за первичные маховые перья, позволяло этим животным более эффективно использовать его при карабканьи или при ковырянии коры. Другие особенности также могут быть связаны с лазающим образом жизни. Первый палец не был противопоставлен остальным, он удерживался на одном уровне с другими пальцами стопы и был направлен вперёд, что, вероятно, помогало при карабканьи по стволам деревьев. Вероятно, эти животные были насекомоядными и вели «более древесный» образ жизни, чем близкие им формы. Возможно, предназначавшиеся к планирующему полёту или парашютированию, маховые перья ювенильных экземпляров имели характерный узор в виде «ёлочки», указывающий на наличие опахала с бородками. При изучении окаменелых останков оказалось, что маховые перья крепились к третьему пальцу; это свидетельствует, что второй и третий палец при жизни были соединены друг с другом мягкой тканью.
Демонстрационное перо Ху Epidexipteryx hui
Время: 155 млн. лет назад.Место находки: Внутренняя Монголия, Китай.Среда обитания: формация Даохугоу (см. выше).Размеры: РК неизвестен; ДТ 30 см (11,8 дюйма); ПД >45 см (1,5 фута).Характерные особенности: Выступающие вперёд зубы. Голова высокая и прямоугольная. Мог иметь длинный третий палец, подобный таковому у Scansoriopteryx.Хвост короткий, имеющий четыре длинных лентообразных пера.Поскольку у них не было истинного пигостиля, хвостовые перья, вероятно, были в большей степени неподвижными по отношению к хвосту и постоянно держались раскрытым веером.Биология:Крылья сильно редуцированы и не имеют первичных маховых перьев, что указывает на потерю способности к планирующему полёту. Вероятно, насекомоядные. Короткий хвост E. hui был сросшимся – состояние, которое, вероятно, развилось независимо от других птиц и связано с укорочением хвоста. Он использовался для поддержки яркого и бросающегося в глаза набора из четырёх очень длинных хвостовых перьев (ETFs – «удлинённые лентообразные хвостовые перья»). Они отличались от современных перьев, но походили на таковые у конфуциусорнитид и энанциорнисов; центральный стержень этих ETF был широким, плоским и не имеющим опахала или бородок; он образовывал единый лентообразный лист.
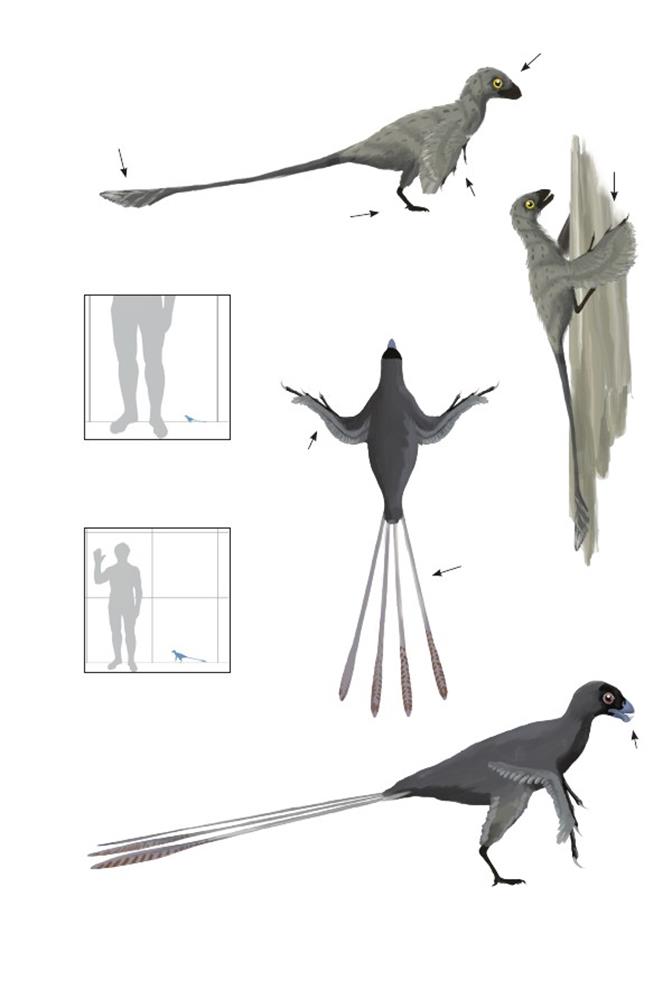
Птица из восточного ЦзисянJixiangornis orientalis
Время: 124,5 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания:Нижняя часть формации Исьянь. Размеры: РК ~1 м (3,3 фута); ДТ 70 см (2,3 фута); ПД неизвестна.Характерные особенности: Голова треугольной формы. Зубы либо отсутствуют, либо очень маленькие и не сохранились в известных экземплярах. Кончик нижней челюсти расширен и выделяется. Тело длинное. Крылья длинные, но детали оперения неизвестны. Первый палец не противопоставлен, но, возможно, был направлен медиально. Хвост умеренно длинный, с рулевыми перьями только лишь на кончике, и короче, чем хвост у сходногоJeholornis prima. Детали строения рулевых перьев неизвестны, но они могли быть ладонеобразными, как у J. prima.Биология: Очень похож на J. prima, отличается более коротким хвостом и очевидным отсутствием зубов.
Первая джехольская птица Jeholornis prima
Время: 120 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания: формация Цзюфотан. Размеры: РК 1 м (3,3 фута); ДТ 60 см (2 фута); ПД 70 см (2,3 фута).Характерные особенности: Голова треугольной формы с несколькими маленькими зубами в верхней и нижней челюсти. «Подбородок» выделяющийся, заострённый у некоторых экземпляров, но округлый у других. Крылья длинные и немного заострённые. Первый палец не противопоставлен. Хвост длинный с коротким листовидным расширением из рулевых перьев на кончике. Рулевые перья узкие, загнуты наружу, с заострёнными не перехлёстывающимися кончиками. Биология: Известно, что питались семенами и, вероятно, собирали их в основном на земле. У маленьких листовидных расширений из рулевых перьев отсутствовали аэродинамические свойства, и они служили, вероятно, для демонстрационного поведения. Jeholornis palmapenis был описан на основе экземпляров той же эпохи, отличавшихся наличием мелких зубов. Однако, возможно, это особенность сохранности материала, и оба они представляют один и тот же вид. Название Jeholornis prima было опубликовано 25 июля 2002 года в еженедельном журнале, тогда как название другого вероятного синонима – Shenzhouraptor sinensis, было опубликовано в ежемесячном журнале без даты. Согласно правилам МКЗН, датой публикации должно считаться 31 июля 2002 года – фактическая дата печати. Несмотря на то, что пресс-релиз, сопровождающий описание Shenzhouraptor, был датирован 23 июля 2002 года, в опубликованных материалах отсутствуют свидетельства того, что пресс-релиз и печатная версия в журнале появились в одно время, поэтому название Jeholornis следует считать более ранним.
Длиннохвостая птица из Яньдана Yandangornis longicaudus
Время: 85 млн. лет назад.Место находки: Чжецзян, Китай.Среда обитания: таншанская группа.Размеры: РК >60 см (2 фута); ДТ 60 см (2 фута); ПД ~65 см (2,1 фута).Характерные особенности:Голова длинная с длинным и заострённым, но массивным беззубым клювом. Крылья длинные. Ноги очень длинные с мощной(ым) мускулатурой/креплением связок на лодыжках. Вероятно, были хорошими бегунами. Когти стоп маленькие и немного изогнутые. Первый палец маленький, расположен высоко на плюсне и не обращён назад. Хвост тонкий и короткий. Рулевые перья ограничены кончиком хвоста. Биология: Вероятно, обитал на побережье мутных рек или озёр, питаясь рыбой и беспозвоночными.
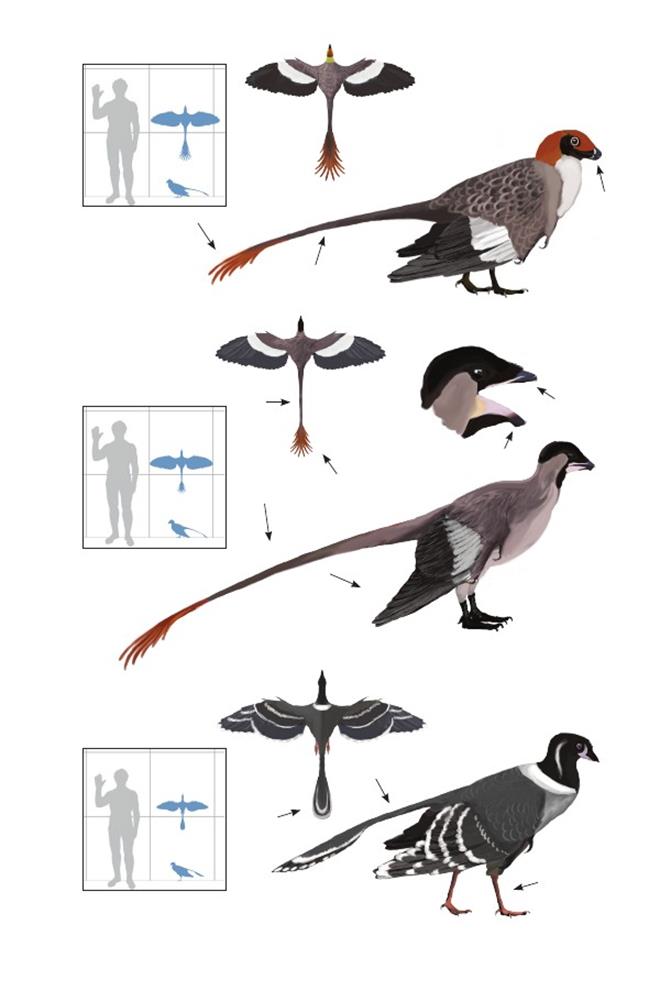
Птица святого Конфуция Confuciusornis sanctus
Время: 124,6-122 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания: формация Исьянь.Размеры: РК 63 см (2 фута); ДТ 25 см (10 дюймов); ПД 50 см (1,6 фута).Характерные особенности: Голова округлая с длинным сильно заострённым клювом. Цвет тела варьирует от чёрного до тёмно-серого с каштановыми или красновато-коричневыми крапинками. Крылья чрезвычайно длинные и заострённые у взрослых особей, у молодых они немного короче. Когти крыльев очень большие, сильно изогнуты и без перьев. Первичные маховые перья белые, вторичные и кроющие перья серые и чёрные. Ноги очень короткие с маленькими стопами и медиальным первым пальцем. Самцы лентохвостые с одной парой очень длинных рулевых перьев. У самок отсутствуют рулевые перья, хвост состоит из крошечных кроющих перьев (которые также присутствуют у самцов). Биология:Молодые самцы имели более длинные костистые хвосты, позвонки которых срастались, а сами хвосты укорачивались по мере взросления. Рулевые перья развиваются у самцов в молодом возрасте или почти в выводковом состоянии; рулевые перья пропорционально длинней у меньших особей. Рост был медленным в течение 1-го года или двух лет, и быстрым от среднего до полного размера. Анатомия крыла радикально отличалась от птичьей; грудь маленькая и неразвитая, грудина имеет только маленький хрящевой киль. Большое отверстие в плечевой кости указывает на уникальную мускулатуру, позволявшую некоторою степень маховых движений, но угол, на который крылья могли бы подниматься над спиной, ограничен анатомией плеча. Вероятно, при карабкании по деревьям с использованием больших когтей крыла, медиальный первый палец помогал перемещаться среди веток. Могли планировать среди деревьев, используя слабые махи. Возможно, пролетали группами над озёрами, где впоследствии многие сохранились. До сих пор наиболее распространённый вид птиц в Исьянь, возможно, они вели колониальный образ жизни, сбиваясь в огромные стаи, некоторые из которых массово гибли во время извержений вулканов и одновременно были захоронены на дне озёр. Молодые особи отсутствуют в крупных скоплениях окаменелостей, что свидетельствует о том, что перед сбиванием в стаи птицы до средних размеров жили поодиночке, или же в более удалённых средах. Рацион этого вида неизвестен.
Птица Конфуция Ду Confuciusornis dui
Время: 124,6 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания: формация Исьянь.Размеры: РК 52 см (1,7 фута); ДТ 17 см (7 дюймов); ПД 40 см (1,3 фута).Характерные особенности: По общей анатомии очень схож с C. sanctus. Отличается меньшим размером, загнутым вверх кончиком клюва и значительно меньшим крылышковым когтем. Биология: Особенная форма клюва явно указывает на уникальный рацион, хотя содержимое желудка неизвестно. Известно, что поздние конфуциусорнитиды питались мелкой рыбой, и это кажется хорошим функциональным применением клюва C. dui.
Предшественник птицы Конфуция Чжена Eoconfuciusornis zhengi
Время: 131 млн. лет назад.Место находки: Хэбей, Китай.Среда обитания:стратон Сычакоу, формация Хуацзиин.Размеры: РК 38 см (1,2 фута); ДТ 17 см (7 дюймов); ПД 40 см (1,3 фута).Характерные особенности: Клюв короткий и заострённый, голова треугольной формы. Крылья заострённые, но широкие и относительно крепкие с длинными вторичными маховыми перьями. Ноги короткие с частично обращённым назад первым пальцем. Лентообразный хвост состоит из одной пары очень длинных рулевых перьев. Биология: Сохранность тёмного меланина в единственном известном экземпляре предполагает очень тёмный, возможно, чёрный окрас. Во многих отношениях похож на Confuciusornis, но гораздо старше хронологически. Также отличается меньшими плечевыми костями (коракоидами) и относительно крепкими позвонками.




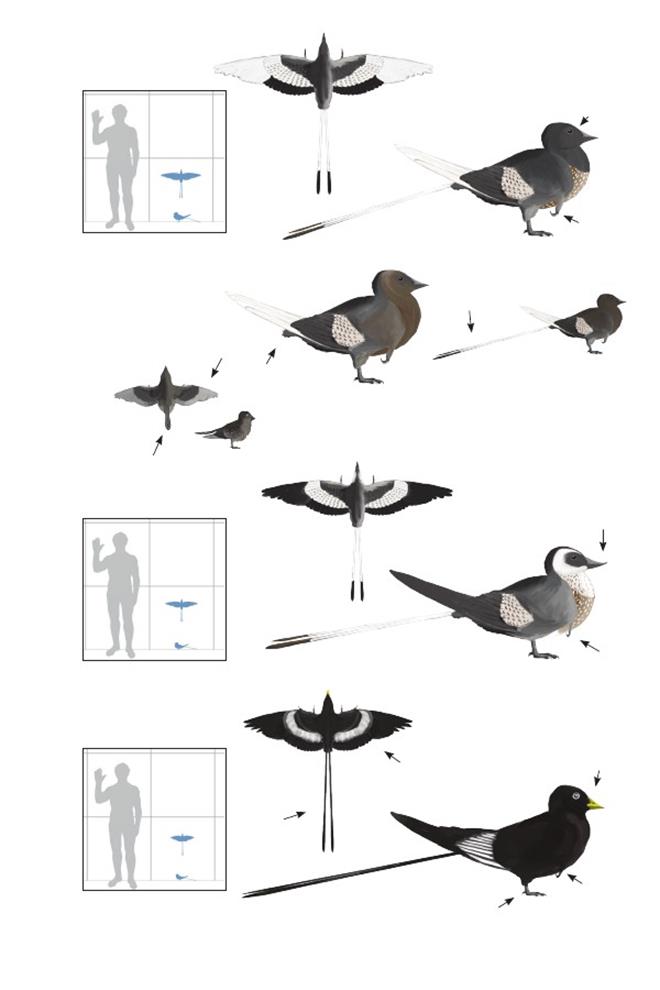
Птица Конфуция из ЦзяньчанConfuciusornis jianchangensis
Время: 120 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания: формация Цзюфотан, умеренно-субтропические болота с преобладанием гинкго и хвойных деревьев, расположенные среди мелководных озёр и стоячих протоков. Размеры: РК неизвестен; ДТ 17 см (7 дюймов); ПД неизвестна.Характерные особенности: Голова маленькая и округлая. Зубы отсутствуют. Клюв короткий. Ноги относительно длинные с короткой плюсной. Крылья и рулевые перья неизвестны. Биология: По общей анатомии очень похож на других конфуциусорнитидов, но в некоторых отношениях более похож на современных птиц (Cau 2010a). Например, короткое туловище с меньшим количеством позвонков, чем у других видов и длинная седалищная кость могут означать, что этот вид –подобный конфуциусорнитидам примитивный эуорнитин или энанциорнитин. Содержимое желудка показывает, что этот вид, по крайней мере, частично питался рыбой.
Птица Великой Стены ХэндаоцзыChangchengornis hengdaoziensis
Время: 122 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания: верхняя часть формации Исьянь (см. выше).Размеры: РК 42 см (1,4 фута); ДТ 20 см (8 дюймов); ПД >30 см (1 фут).Характерные особенности:Очень маленький. Клюв длинный и крючковатый. Гребень отчётливо округлый. Крылья короткие и относительно широкие. Ноги короткие с частично противопоставленным первым пальцем. Лентообразный хвост с одной парой рулевых перьев меньшего размера, чем у родственных видов.Биология: Возможно, вели наполовину древесный образ жизни, но имели меньшие размеры, неспециализированные крылья, указывающие на плохой полёт. Вероятно, кормились в основном на земле.
SAPE(аббревиатура от англ.: Society of Avian Paleontology and Evolution (Сообщество палеонтологии и эволюции птиц) птица Чаояна Sapeornis chaoyangensis
Время: 120 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания: формация Цзюфотан.Размеры: РК 1,4 м (4,6 фута); ДТ 45 см (1,5 фута); ПД неизвестна.Характерные особенности:Голова высокая и округлая с узкой мордой. Глаза большие. Выступающие вперёд зубы ограничены кончиком верхней челюсти. Нижняя челюсть беззубая. Крылья очень длинные с большими когтями и длинными первичными маховыми перьями (третье первичное перо самое длинное, первое – самое короткое). Хвост короткий. Ноги короткие и крепкие. Пальцы стоп очень длинные с сильно загнутыми когтями. Первый палец обращён назад. Биология: Окаменелости, найденные в наземных и озёрных отложениях, указывают на широкое распространение (возможно, в первую очередь, населяли кроны деревьев). Ловко садясь на ветки, они могли карабкаться вверх по стволам деревьев, используя большие когти крыльев, чтобы спастись от хищников, и проводили большую часть времени, карабкаясь среди высоких ветвей. Неспособные к сильному машущему полёту, широкие крылья могли использоваться для длительных скользящих/парящих полётов между деревьями. Были растительноядными или всеядными, возможно, ели листья и семена, дополняя свой рацион фруктами и беспозвоночными. Единственный хорошо известный представитель Omnivoropterygiformes – «всеядные крылья» – причудливой ранней группы короткохвостых птиц, в некотором роде похожей на ценагнатиформов. Были описаны некоторые другие виды, однако все они, вероятно, представляют этапы роста одного этого вида.
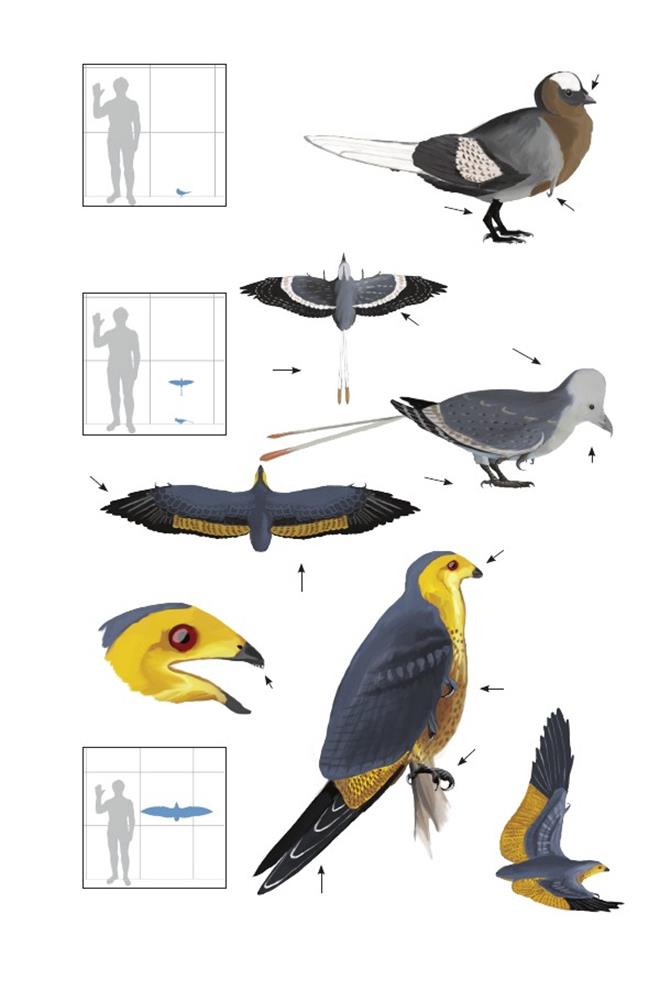
Базальные энанциорнитины
Наиболее разнообразной и успешной группой мезозойских птиц были энанциорнитины («противоптицы»). Они получили своё название из-за того, что их плечевой сустав крепится противоположным образом по сравнению с креплением плечевого сустава эуорнитинов («истинных птиц»). У современных птиц лопатка имеет выступ, входящий во впадину плечевого пояса (коракоида). У энанциорнитин наоборот, впадина имеется на лопатке. Это может свидетельствовать о независимых путях эволюции устойчивого машущего полёта.
Как представляется, большинство энанциорнитин, были наземными птицами; они населяли покрытые лесом материковые части суши и, соответственно, были приспособлены к жизни на деревьях, хотя некоторые виды, возможно, также свободно кормились на земле или ловили рыбу с берега. Энанциорнитины имели резко обращённые назад первые пальцы задних конечностей. Известно, что некоторые виды гнездились большими колониями на земле вблизи рек, озёр и других водоёмов (Dyke & al, 2011).
Как и большинство мезозойских птиц, энанциорнитины могли обладать слегка отличающимся характером роста от типичных современных птиц.Результаты исследований колец роста их окаменевших костей указывают, что, хотя после вылупления они быстро росли, их рост значительно замедлялся до достижения полного размера (Cambra-Moo & al., 2006).Как было установлено, у некоторых видов энанциорнитин быстро росли крылья, и они, вероятно, были способны летать уже в относительно молодом возрасте.У этих видов продолжительность родительской заботы могла быть сравнительно недолгой.Поскольку в течение первого года жизни, в отличие от большинства современных птиц, они не достигали полного размера, молодые энанциорнитины могли играть другие роли и занимать другие экологические ниши, нежели их родители, меняя по мере взросления свой рацион и экологию, как это предполагается для более примитивных птиц и других тероподов.
Из-за их уникальной анатомии и повсеместного распространения, окаменевшие остатки энанциорнитин легко идентифицируются, но многие из них были описаны на основе крайне фрагментарных ископаемых остатков, часто лишь по маленьким фрагментам кости. Поэтому многие виды, хотя и, несомненно, уникальные, имеют небольшое научное значение. Кроме того, родственные связи энанциорнитин изучены очень плохо; достаточно обоснованным является существование лишь одной или двух групп. Фрагментарный характер остатков большинства видов, невозможность использовать филогенетический брекетинг из-за плохо изученных родственных связей и полнейшее разнообразие форм хорошо известных видов делает невозможной достоверную реконструкцию большинства энанциорнитин. По этой причине ниже представлены только хорошо изученные представители, в большинстве своём те, для которых известен черепной материал.
Примитивное перо из Фэннина Protopteryx fengningensis
Время: 131 млн. лет назад.Место находки: Хэбэй, Китай. Среда обитания: стратон Сычакоу, формация Хуацзиинь. Леса умеренной климатической зоны с преобладанием озёр. Размеры: РК 33 см (1 фут); ДТ 16 см (6 дюймов); ПД >23 см (9 дюймов). Характерные особенности: Голова круглая с несколько заострённой мордой, зубы ограничены на кончиках верхней и нижней челюсти. Крылья относительно короткие и широкие (средняя длина первичного махового пера 45 мм, самое длинное маховое перо – 95 мм). Одна пара длинных, тонких лентовидных рулевых перьев.Биология: Наиболее ранняя из известных птиц, определённо способная к активному полёту. Короткие широкие крылья, должно быть, были приспособлены к маневрированию в условиях густого леса, хотя отсутствие веерообразного хвоста и примитивный длинный палец крылышка, вероятно, делали её полёт более неуклюжим по сравнению с современными древесными птицами. Летательный аппарат хорошо развит, грудина имеет киль, но крылья сохраняют раздельные костные пальцы и маленькие когти.
Стройный сходный с протоптериксомParaprotopteryx gracilis
Время: 124,6 млн. лет назад.Место находки: Ляонин, Китай.Среда обитания:формация Нижний Исянь. Лесные массивы из хвойных и гинкго умеренной климатической зоны среди системы озёр, подпитываемых ручьями и притоком с близлежащей цепи активных вулканов. Размеры: (ювенильная особь) РК >22 см (9 дюймов); ДТ >11 см (4 дюйма); ПД >20 см (8 дюймов). Характерные особенности: Известен только по остаткам почти взрослой особи. Крылья короткие (средняя длина первичного махового пера 40 мм). Кисть срослась у основания. Палец крылышка с маленьким когтем. Коготь большого пальца меньше когтя пальца крылышка, а коготь третьего пальца рудиментарный. Ноги относительно короткие, со стопами, имеющими сильные когти и обращённый назад первый палец. Хвост короткий с четырьмя относительно короткими и тонкими лентовидными рулевыми перьями. Каждое лентовидное перо оканчивается овальным расширением.Биология: Анатомия крыла характеризуется отсутствием прокоракоидной структуры, обнаруженной у близкородственного вида Protopteryx, что говорит о несколько худшей приспособленности к полёту.Как и другие примитивные энанциорнитины, для карабкания по деревьям, вероятно, пользовались в основном большими когтями крыльев.
Дата: 2019-07-24, просмотров: 689.